
Nanoarchaeum equitans is a species of marine archaea that was discovered in 2002 in a hydrothermal vent off the coast of Iceland on the Kolbeinsey Ridge by Karl Stetter. It has been proposed as the first species in a new phylum, and is the only species within the genus Nanoarchaeum. Strains of this microbe were also found on the Sub-polar Mid Oceanic Ridge, and in the Obsidian Pool in Yellowstone National Park. Since it grows in temperatures approaching boiling, at about 80 °C (176 °F), it is considered to be a thermophile. It grows best in environments with a pH of 6, and a salinity concentration of 2%. Nanoarchaeum appears to be an obligate symbiont on the archaeon Ignicoccus; it must be in contact with the host organism to survive. Nanoarchaeum equitans cannot synthesize lipids but obtains them from its host. Its cells are only 400 nm in diameter, making it the smallest known living organism, and the smallest known archaeon.
Methanogens are anaerobic archaea that produce methane as a byproduct of their energy metabolism, i.e., catabolism. Methane production, or methanogenesis, is the only biochemical pathway for ATP generation in methanogens. All known methanogens belong exclusively to the domain Archaea, although some bacteria, plants, and animal cells are also known to produce methane. However, the biochemical pathway for methane production in these organisms differs from that in methanogens and does not contribute to ATP formation. Methanogens belong to various phyla within the domain Archaea. Previous studies placed all known methanogens into the superphylum Euryarchaeota. However, recent phylogenomic data have led to their reclassification into several different phyla. Methanogens are common in various anoxic environments, such as marine and freshwater sediments, wetlands, the digestive tracts of animals, wastewater treatment plants, rice paddy soil, and landfills. While some methanogens are extremophiles, such as Methanopyrus kandleri, which grows between 84 and 110°C, or Methanonatronarchaeum thermophilum, which grows at a pH range of 8.2 to 10.2 and a Na+ concentration of 3 to 4.8 M, most of the isolates are mesophilic and grow around neutral pH.

Methanosarcina is a genus of euryarchaeote archaea that produce methane. These single-celled organisms are known as anaerobic methanogens that produce methane using all three metabolic pathways for methanogenesis. They live in diverse environments where they can remain safe from the effects of oxygen, whether on the earth's surface, in groundwater, in deep sea vents, and in animal digestive tracts. Methanosarcina grow in colonies.
Microbial metabolism is the means by which a microbe obtains the energy and nutrients it needs to live and reproduce. Microbes use many different types of metabolic strategies and species can often be differentiated from each other based on metabolic characteristics. The specific metabolic properties of a microbe are the major factors in determining that microbe's ecological niche, and often allow for that microbe to be useful in industrial processes or responsible for biogeochemical cycles.
In biology, syntrophy, syntrophism, or cross-feeding is the cooperative interaction between at least two microbial species to degrade a single substrate. This type of biological interaction typically involves the transfer of one or more metabolic intermediates between two or more metabolically diverse microbial species living in close proximity to each other. Thus, syntrophy can be considered an obligatory interdependency and a mutualistic metabolism between different microbial species, wherein the growth of one partner depends on the nutrients, growth factors, or substrates provided by the other(s).
Methanosarcinales is an order of Archaea in the class Methanomicrobia, phylum Methanobacteriota. The order Methanosarcinales contains both methanogenic and methanotrophic lineages, although the latter so far have so far no pure culture representatives. Methanotrophic lineages of the order Methanosarcinales were initially abbreviated as ANME to distinguich from aerobic methanotrophic bacteria. Currently, those lineages receive their own names such as Ca. Methanoperedens, Ca. Methanocomedens (ANME-2a), Ca.Methanomarinus (ANME-2b), Ca. Methanogaster (ANME-2c), Ca. Methanovorans (ANME-3). The order contains archaeon with one of the largest genome, Methanosarcina acetivorans C2A, genome size 5,75 Mbp.

The Wood–Ljungdahl pathway is a set of biochemical reactions used by some bacteria. It is also known as the reductive acetyl-coenzyme A (acetyl-CoA) pathway. This pathway enables these organisms to use hydrogen as an electron donor, and carbon dioxide as an electron acceptor and as a building block for biosynthesis.
In taxonomy, Thermococcus is a genus of thermophilic Archaea in the family the Thermococcaceae.

Nitrosopumilus is a genus of archaea. The type species, Nitrosopumilus maritimus, is an extremely common archaeon living in seawater. It is the first member of the Group 1a Nitrososphaerota to be isolated in pure culture. Gene sequences suggest that the Group 1a Nitrososphaerota are ubiquitous with the oligotrophic surface ocean and can be found in most non-coastal marine waters around the planet. It is one of the smallest living organisms at 0.2 micrometers in diameter. Cells in the species N. maritimus are shaped like peanuts and can be found both as individuals and in loose aggregates. They oxidize ammonia to nitrite and members of N. maritimus can oxidize ammonia at levels as low as 10 nanomolar, near the limit to sustain its life. Archaea in the species N. maritimus live in oxygen-depleted habitats. Oxygen needed for ammonia oxidation might be produced by novel pathway which generates oxygen and dinitrogen. N. maritimus is thus among organisms which are able to produce oxygen in dark.

Methanobacterium is a genus of the Methanobacteria class in the Archaea kingdom, which produce methane as a metabolic byproduct. Despite the name, this genus belongs not to the bacterial domain but the archaeal domain. Methanobacterium are nonmotile and live without oxygen, which is toxic to them, and they only inhabit anoxic environments.
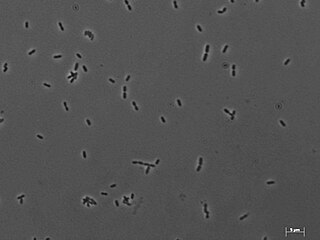
Methanobrevibacter smithii is the predominant methanogenic archaeon in the microbiota of the human gut. M. smithii has a coccobacillus shape. It plays an important role in the efficient digestion of polysaccharides (complex sugars) by consuming the end products of bacterial fermentation (H2, acetate, formate to some extant). M. smithii is a hydrogenotrophic methanogen that utilizes hydrogen by combining it with carbon dioxide to form methane. The removal of hydrogen by M. smithii is thought to allow an increase in the extraction of energy from nutrients by shifting bacterial fermentation to more oxidized end products.

Archaea is a domain of single-celled organisms. These microorganisms lack cell nuclei and are therefore prokaryotic. Archaea were initially classified as bacteria, receiving the name archaebacteria, but this term has fallen out of use.
The Nitrososphaerota are a phylum of the Archaea proposed in 2008 after the genome of Cenarchaeum symbiosum was sequenced and found to differ significantly from other members of the hyperthermophilic phylum Thermoproteota. Three described species in addition to C. symbiosum are Nitrosopumilus maritimus, Nitrososphaera viennensis, and Nitrososphaera gargensis. The phylum was proposed in 2008 based on phylogenetic data, such as the sequences of these organisms' ribosomal RNA genes, and the presence of a form of type I topoisomerase that was previously thought to be unique to the eukaryotes. This assignment was confirmed by further analysis published in 2010 that examined the genomes of the ammonia-oxidizing archaea Nitrosopumilus maritimus and Nitrososphaera gargensis, concluding that these species form a distinct lineage that includes Cenarchaeum symbiosum. The lipid crenarchaeol has been found only in Nitrososphaerota, making it a potential biomarker for the phylum. Most organisms of this lineage thus far identified are chemolithoautotrophic ammonia-oxidizers and may play important roles in biogeochemical cycles, such as the nitrogen cycle and the carbon cycle. Metagenomic sequencing indicates that they constitute ~1% of the sea surface metagenome across many sites.
Methanococcoides burtonii is a methylotrophic methanogenic archaeon first isolated from Ace Lake, Antarctica. Its type strain is DSM 6242.

Methanococcus maripaludis is a species of methanogenic archaea found in marine environments, predominantly salt marshes. M. maripaludis is a non-pathogenic, gram-negative, weakly motile, non-spore-forming, and strictly anaerobic mesophile. It is classified as a chemolithoautotroph. This archaeon has a pleomorphic coccoid-rod shape of 1.2 by 1.6 μm, in average size, and has many unique metabolic processes that aid in survival. M. maripaludis also has a sequenced genome consisting of around 1.7 Mbp with over 1,700 identified protein-coding genes. In ideal conditions, M. maripaludis grows quickly and can double every two hours.
Methanosarcina barkeri is the most fundamental species of the genus Methanosarcina, and their properties apply generally to the genus Methanosarcina. Methanosarcina barkeri can produce methane anaerobically through different metabolic pathways. M. barkeri can subsume a variety of molecules for ATP production, including methanol, acetate, methylamines, and different forms of hydrogen and carbon dioxide. Although it is a slow developer and is sensitive to change in environmental conditions, M. barkeri is able to grow in a variety of different substrates, adding to its appeal for genetic analysis. Additionally, M. barkeri is the first organism in which the amino acid pyrrolysine was found. Furthermore, two strains of M. barkeri, M. b. Fusaro and M. b. MS have been identified to possess an F-type ATPase along with an A-type ATPase.
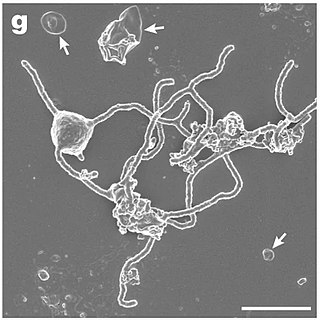
Lokiarchaeota is a proposed phylum of the Archaea. The phylum includes all members of the group previously named Deep Sea Archaeal Group, also known as Marine Benthic Group B. Lokiarchaeota is part of the superphylum Asgard containing the phyla: Lokiarchaeota, Thorarchaeota, Odinarchaeota, Heimdallarchaeota, and Helarchaeota. A phylogenetic analysis disclosed a monophyletic grouping of the Lokiarchaeota with the eukaryotes. The analysis revealed several genes with cell membrane-related functions. The presence of such genes support the hypothesis of an archaeal host for the emergence of the eukaryotes; the eocyte-like scenarios.
"Candidatus Thorarchaeota", or simply Thorarchaeota, is a phylum within the superphylum Asgard archaea. The Asgard superphylum represents the closest prokaryotic relatives of eukaryotes. Since there is such a close relation between the two different domains, it provides further evidence to the two-domain tree of life theory which states that eukaryotes branched from the archaeal domain. Asgard archaea are single cell marine microbes that contain branch like appendages and have genes that are similar to eukarya. The asgard archaea superphylum is composed of Thorarchaeota, Lokiarchaeota, Odinarchaeota, and Heimdallarchaeota. Thorarchaeota were first identified from the sulfate-methane transition zone in tidewater sediments. Thorarcheota are widely distributed in marine and freshwater sediments.

The hydrothermal vent microbial community includes all unicellular organisms that live and reproduce in a chemically distinct area around hydrothermal vents. These include organisms in the microbial mat, free floating cells, or bacteria in an endosymbiotic relationship with animals. Chemolithoautotrophic bacteria derive nutrients and energy from the geological activity at Hydrothermal vents to fix carbon into organic forms. Viruses are also a part of the hydrothermal vent microbial community and their influence on the microbial ecology in these ecosystems is a burgeoning field of research.
The deep biosphere is the part of the biosphere that resides below the first few meters of the surface. It extends down below 10 kilometers below the continental surface and 21 kilometers below the sea surface, at temperatures that may reach beyond 120 °C (248 °F) which is comparable to the maximum temperature where a metabolically active organism has been found. It includes all three domains of life and the genetic diversity rivals that on the surface.
