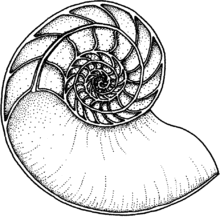
Ammonoids are extinct spiral shelled cephalopods comprising the subclass Ammonoidea. They are more closely related to living coleoids than they are to shelled nautiloids. The earliest ammonoids appeared during the Devonian, with the last species vanishing during or soon after the Cretaceous–Paleogene extinction event. They are often called ammonites, which is most frequently used for members of the order Ammonitida, the only living group of ammonoids from the Jurassic up until their extinction.

Belemnoids are an extinct group of marine cephalopod, very similar in many ways to the modern squid. Like them, the belemnoids possessed an ink sac, but, unlike the squid, they possessed ten arms of roughly equal length, and no tentacles. The name "belemnoid" comes from the Greek word βέλεμνον, belemnon meaning "a dart or arrow" and the Greek word είδος, eidos meaning "form".

Nautiloids are a group of marine cephalopods (Mollusca) which originated in the Late Cambrian and are represented today by the living Nautilus and Allonautilus. Fossil nautiloids are diverse and species rich, with over 2,500 recorded species. They flourished during the early Paleozoic era, when they constituted the main predatory animals. Early in their evolution, nautiloids developed an extraordinary diversity of shell shapes, including coiled morphologies and giant straight-shelled forms (orthocones). No orthoconic and only a handful of coiled species, the nautiluses, survive to the present day.
Rayonnoceras is a genus of extinct cephalopods that lived around 325 million years ago during the Carboniferous. Although they resemble earlier actinocerids they are now thought to belong to the Pseudorthocerida

Endocerida is an extinct nautiloid order, a group of cephalopods from the Lower Paleozoic with cone-like deposits in their siphuncle. Endocerida was a diverse group of cephalopods that lived from the Early Ordovician possibly to the Late Silurian. Their shells were variable in form. Some were straight (orthoconic), others curved (cyrtoconic); some were long (longiconic), others short (breviconic). Some long-shelled forms like Endoceras attained shell lengths close to 6 metres (20 ft). The related Cameroceras is anecdotally reported to have reached lengths approaching 9 metres (30 ft), but these claims are problematic. The overwhelming majority of endocerids and nautiloids in general are much smaller, usually less than a meter long when fully grown.

Orthocerida, also known as the Michelinocerida, is an order of extinct orthoceratoid cephalopods that lived from the Early Ordovician possibly to the Late Triassic. A fossil found in the Caucasus suggests they may even have survived until the Early Cretaceous, and the Eocene fossil Antarcticeras is sometimes considered a descendant of the orthocerids although this is disputed. They were most common however from the Ordovician to the Devonian.

The Bactritida are a small order of more or less straight-shelled (orthoconic) cephalopods that first appeared during the Emsian stage of the Devonian period with questionable origins in the Pragian stage before 409 million years ago, and persisted until the Carnian pluvial event in the upper middle Carnian stage of the Triassic period. They are considered ancestors of the ammonoids, as well as of the coleoids.
Cameroceras is an extinct genus of endocerid cephalopod which lived in equatorial oceans during the entire Ordovician period. Like other endocerids, it was an orthocone, meaning that its shell was fairly straight and pointed. It was particularly abundant and widespread in the Late Ordovician, inhabiting the shallow tropical seas in and around Laurentia, Baltica and Siberia.
Discosorida are an order of cephalopods that lived from the beginning of the Middle Ordovician, through the Silurian, and into the Devonian. Discosorids are unique in the structure and formation of the siphuncle, the tube that runs through and connects the camerae (chambers) in cephalopods, which unlike those in other orders is zoned longitudinally along the segments rather than laterally. Siphuncle structure indicated that the Discosorida evolved directly from the Plectronoceratida rather than through the more developed Ellesmerocerida, as did the other orders. Finally and most diagnostic, discosorids developed a reinforcing, grommet-like structure in the septal opening of the siphuncle known as the bullette, formed by a thickening of the connecting ring as it draped around the folded back septal neck.
The Ellesmerocerida is an order of primitive cephalopods belonging to the subclass Nautiloidea with a widespread distribution that lived during the Late Cambrian and Ordovician.
Baltoceratidae is an extinct family of orthoconic cephalopods belonging to the subclass Nautiloidea endemic to what would be Asia, Australia, Europe, North America, and South America during the Ordovician living from about 480–460 mya, existing for approximately 20 million years.
Sactoceras is an extinct nautiloid cephalopod that lived during the Ordovician and Silurian in what would become North America, Europe, and Asia.

Orthoceratoidea is a major subclass of nautiloid cephalopods. Members of this subclass usually have orthoconic (straight) to slightly cyrtoconic (curved) shells, and central to subcentral siphuncles which may bear internal deposits. Orthoceratoids are also characterized by dorsomyarian muscle scars, extensive cameral deposits, and calciosiphonate connecting rings with a porous and calcitic inner layer.

The Armenoceratidae are a family of early Paleozoic nautiloid cephalopods belonging to the order Actinocerida.
Macroloxoceras is a large pseuorthocerid from the upper Devonian of Central Colorado and Southern New Mexico with features resembling those found in actinocerids. Pseudorthocerids and actinocerids are extinct nautiloid cephalopods, generally with long straight shells and expanded siphuncle segments filled with organic deposits.

Endoceratidae is a family of large to very large straight shelled nautiloid cephalopods belonging to the order Endocerida that lived during the Middle and Late Ordovician. They include the largest known Paleozoic invertebrates, represented by Endoceras and Cameroceras.
Dolorthoceras is a nautiloid cephalopod from the upper Paleozoic found in Lower Devonian to Lower Permian strata in North America, Europe, Asia, and Australia.
Cyrtocerinida is an order of Ordovician nautiloid cephalopods. The order includes the families Cyrtocerinidae and Eothinoceratidae, as well as the genera Bathmoceras and Rummoceras.

Multiceratoidea is a major subclass or superorder of Paleozoic nautiloid cephalopods. Members of this group can be characterized by nautilosiphonate connecting rings, with an organic inner layer and outer layer of calcitic spherules and blades, similar to the modern nautilus. The earliest-diverging multiceratoids have oncomyarian muscle scars, though several orders trend towards a ventromyarian condition. Multiceratoid shells are generally short and curled, with a relatively small aperture (opening). Cameral deposits are never found among the multiceratoids, though several orders are known to bear endosiphuncular deposits within their siphuncles.
Bisonocerida is an order of Ordovician to Silurian nautiloid cephalopods. Members of this order were originally placed in the order Endocerida, but later investigation argued that this broad usage of Endocerida was a polyphyletic assemblage encompassing two different groups of independent origin. Bisonocerida was differentiated from Endocerida in 2012 in order to resolve this issue.