Chromatophores are largely responsible for generating skin and eye colour in ectothermic animals and are generated in the neural crest during embryonic development. Mature chromatophores are grouped into subclasses based on their colour under white light: xanthophores (yellow), erythrophores (red), iridophores (reflective / iridescent), leucophores (white), melanophores (black/brown), and cyanophores (blue). While most chromatophores contain pigments that absorb specific wavelengths of light, the color of leucophores and iridophores is produced by their respective scattering and optical interference properties.
7x speed timelapse video of fish melanophores responding to 200μM adrenaline
Some species can rapidly change colour through mechanisms that translocate pigment and reorient reflective plates within chromatophores. This process, often used as a type of camouflage, is called physiological colour change or metachrosis.[1] Cephalopods, such as the octopus, have complex chromatophore organs controlled by muscles to achieve this, whereas vertebrates such as chameleons generate a similar effect by cell signalling. Such signals can be hormones or neurotransmitters and may be initiated by changes in mood, temperature, stress or visible changes in the local environment.[citation needed] Chromatophores are studied by scientists to understand human disease and as a tool in drug discovery.
These animals also escape detection by a very extraordinary, chameleon-like power of changing their colour. They appear to vary their tints according to the nature of the ground over which they pass: when in deep water, their general shade was brownish purple, but when placed on the land, or in shallow water, this dark tint changed into one of a yellowish green. The colour, examined more carefully, was a French grey, with numerous minute spots of bright yellow: the former of these varied in intensity; the latter entirely disappeared and appeared again by turns. These changes were effected in such a manner that clouds, varying in tint between a hyacinth red and a chestnut-brown, were continually passing over the body. Any part, being subjected to a slight shock of galvanism, became almost black: a similar effect, but in a less degree, was produced by scratching the skin with a needle. These clouds, or blushes as they may be called, are said to be produced by the alternate expansion and contraction of minute vesicles containing variously coloured fluids.
Classification of chromatophore
A veiled chameleon, Chamaeleo calyptratus. Structural green and blue colours are generated by overlaying chromatophore types to reflect filtered light.
The term chromatophore was adopted (following Sangiovanni's chromoforo) as the name for pigment-bearing cells derived from the neural crest of cold-blooded vertebrates and cephalopods. The word itself comes from the Greek words chrōma (χρῶμα) meaning "colour," and phoros (φόρος) meaning "bearing". In contrast, the word chromatocyte (kytos (κύτος) meaning "cell") was adopted for the cells responsible for colour found in birds and mammals. Only one such cell type, the melanocyte, has been identified in these animals.
It was only in the 1960s that chromatophores were well enough understood to enable them to be classified based on their appearance. This classification system persists to this day, even though the biochemistry of the pigments may be more useful to a scientific understanding of how the cells function.[5]
Colour-producing molecules fall into two distinct classes: biochromes and structural colours or "schemochromes".[6] The biochromes include true pigments, such as carotenoids and pteridines. These pigments selectively absorb parts of the visible light spectrum that makes up white light while permitting other wavelengths to reach the eye of the observer. Structural colours are produced by various combinations of diffraction, reflection or scattering of light from structures with a scale around a quarter of the wavelength of light. Many such structures interfere with some wavelengths (colours) of light and transmit others, simply because of their scale, so they often produce iridescence by creating different colours when seen from different directions.[citation needed]
Whereas all chromatophores contain pigments or reflecting structures (except when there has been a mutation, as in albinism), not all pigment-containing cells are chromatophores. Haem, for example, is a biochrome responsible for the red appearance of blood. It is found primarily in red blood cells (erythrocytes), which are generated in bone marrow throughout the life of an organism, rather than being formed during embryological development. Therefore, erythrocytes are not classified as chromatophores.[citation needed]
Xanthophores and erythrophores
Chromatophores that contain large amounts of yellow pteridine pigments are named xanthophores; those with mainly red/orange carotenoids are termed erythrophores.[5] However, vesicles containing pteridine and carotenoids are sometimes found in the same cell, in which case the overall colour depends on the ratio of red and yellow pigments.[7] Therefore, the distinction between these chromatophore types is not always clear.
Most chromatophores can generate pteridines from guanosine triphosphate, but xanthophores appear to have supplemental biochemical pathways enabling them to accumulate yellow pigment. In contrast, carotenoids are metabolised and transported to erythrophores. This was first demonstrated by rearing normally green frogs on a diet of carotene-restricted crickets. The absence of carotene in the frogs' diet meant that the red/orange carotenoid colour 'filter' was not present in their erythrophores. This made the frogs appear blue instead of green.[8]
Iridophores and leucophores
Leucophore layer composition
Iridophores, sometimes also called guanophores, are chromatophores that reflect light using plates of crystalline chemochromes made from guanine.[9] When illuminated they generate iridescent colours because of the constructive interference of light. Fish iridophores are typically stacked guanine plates separated by layers of cytoplasm to form microscopic, one-dimensional, Bragg mirrors. Both the orientation and the optical thickness of the chemochrome determines the nature of the colour observed.[10] By using biochromes as coloured filters, iridophores create an optical effect known as Tyndall or Rayleigh scattering, producing bright-blue or -green colours.[11]
A related type of chromatophore, the leucophore, is found in some fish, in particular in the tapetum lucidum. Like iridophores, they utilize crystalline purines (often guanine) to reflect light. Unlike iridophores, leucophores have more organized crystals that reduce diffraction. Given a source of white light, they produce a white shine. As with xanthophores and erythrophores, in fish the distinction between iridophores and leucophores is not always obvious, but, in general, iridophores are considered to generate iridescent or metallic colours, whereas leucophores produce reflective white hues.[11]
Melanophores
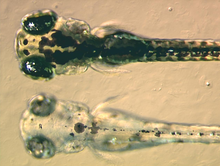
At the bottom a mutant zebrafish larva that fails to synthesise melanin in its melanophores, at the top a non-mutant, wildtype larva
Melanophores contain eumelanin, a type of melanin, that appears black or dark-brown because of its light absorbing qualities. It is packaged in vesicles called melanosomes and distributed throughout the cell. Eumelanin is generated from tyrosine in a series of catalysed chemical reactions. It is a complex chemical containing units of dihydroxyindole and dihydroxyindole-2-carboxylic acid with some pyrrole rings.[12] The key enzyme in melanin synthesis is tyrosinase. When this protein is defective, no melanin can be generated resulting in certain types of albinism. In some amphibian species there are other pigments packaged alongside eumelanin. For example, a novel deep (wine) red-colour pigment was identified in the melanophores of phyllomedusine frogs.[13] Some species of anole lizards, such as the Anolis grahami, use melanocytes in response to certain signals and hormonal changes, and is capable of becoming colors ranging from bright blue, brown, and black. This was subsequently identified as pterorhodin, a pteridine dimer that accumulates around eumelanin core, and it is also present in a variety of tree frog species from Australia and Papua New Guinea. While it is likely that other lesser-studied species have complex melanophore pigments, it is nevertheless true that the majority of melanophores studied to date do contain eumelanin exclusively.[14]
Humans have only one class of pigment cell, the mammalian equivalent of melanophores, to generate skin, hair, and eye colour. For this reason, and because the large number and contrasting colour of the cells usually make them very easy to visualise, melanophores are by far the most widely studied chromatophore. However, there are differences between the biology of melanophores and that of melanocytes. In addition to eumelanin, melanocytes can generate a yellow/red pigment called phaeomelanin.[citation needed]
The purple-striped dottyback, Pseudochromis diadema, generates its violet stripe with an unusual type of chromatophore.
Cyanophores
Nearly all the vibrant blues in animals and plants are created by structural coloration rather than by pigments. However, some types of Synchiropus splendidus do possess vesicles of a cyan biochrome of unknown chemical structure in cells named cyanophores.[11] Although they appear unusual in their limited taxonomic range, there may be cyanophores (as well as further unusual chromatophore types) in other fish and amphibians. For example, brightly coloured chromatophores with undefined pigments are found in both poison dart frogs and glass frogs,[15] and atypical dichromatic chromatophores, named erythro-iridophores have been described in Pseudochromis diadema.[16]
Pigment translocation
Fish and frog melanophores are cells that can change colour by dispersing or aggregating pigment-containing bodies.
Many species are able to translocate the pigment inside their chromatophores, resulting in an apparent change in body colour. This process, known as physiological colour change, is most widely studied in melanophores, since melanin is the darkest and most visible pigment. In most species with a relatively thin dermis, the dermal melanophores tend to be flat and cover a large surface area. However, in animals with thick dermal layers, such as adult reptiles, dermal melanophores often form three-dimensional units with other chromatophores. These dermal chromatophore units (DCU) consist of an uppermost xanthophore or erythrophore layer, then an iridophore layer, and finally a basket-like melanophore layer with processes covering the iridophores.[17]
Both types of melanophore are important in physiological colour change. Flat dermal melanophores often overlay other chromatophores, so when the pigment is dispersed throughout the cell the skin appears dark. When the pigment is aggregated toward the centre of the cell, the pigments in other chromatophores are exposed to light and the skin takes on their hue. Likewise, after melanin aggregation in DCUs, the skin appears green through xanthophore (yellow) filtering of scattered light from the iridophore layer. On the dispersion of melanin, the light is no longer scattered and the skin appears dark. As the other biochromatic chromatophores are also capable of pigment translocation, animals with multiple chromatophore types can generate a spectacular array of skin colours by making good use of the divisional effect.[18][19]
A single zebrafish melanophore imaged by time-lapse photography during pigment aggregation
The control and mechanics of rapid pigment translocation has been well studied in a number of different species, in particular amphibians and teleost fish.[11][20] It has been demonstrated that the process can be under hormonal or neuronal control or both and for many species of bony fishes it is known that chromatophores can respond directly to environmental stimuli like visible light, UV-radiation, temperature, pH, chemicals, etc.[21] Neurochemicals that are known to translocate pigment include noradrenaline, through its receptor on the surface on melanophores.[22] The primary hormones involved in regulating translocation appear to be the melanocortins, melatonin, and melanin-concentrating hormone (MCH), that are produced mainly in the pituitary, pineal gland, and hypothalamus, respectively. These hormones may also be generated in a paracrine fashion by cells in the skin. At the surface of the melanophore, the hormones have been shown to activate specific G-protein-coupled receptors that, in turn, transduce the signal into the cell. Melanocortins result in the dispersion of pigment, while melatonin and MCH results in aggregation.[23]
Most fish, reptiles and amphibians undergo a limited physiological colour change in response to a change in environment. This type of camouflage, known as background adaptation, most commonly appears as a slight darkening or lightening of skin tone to approximately mimic the hue of the immediate environment. It has been demonstrated that the background adaptation process is vision-dependent (it appears the animal needs to be able to see the environment to adapt to it),[32] and that melanin translocation in melanophores is the major factor in colour change.[23] Some animals, such as chameleons and anoles, have a highly developed background adaptation response capable of generating a number of different colours very rapidly.[33] They have adapted the capability to change colour in response to temperature, mood, stress levels, and social cues, rather than to simply mimic their environment.
Development
Cross-section of a developing vertebrate trunk showing the dorsolateral (red) and ventromedial (blue) routes of chromatoblast migration
During vertebrate embryonic development, chromatophores are one of a number of cell types generated in the neural crest, a paired strip of cells arising at the margins of the neural tube. These cells have the ability to migrate long distances, allowing chromatophores to populate many organs of the body, including the skin, eye, ear, and brain. Fish melanophores and iridophores have been found to contain the smooth muscle regulatory proteins [calponin] and caldesmon.[34] Leaving the neural crest in waves, chromatophores take either a dorsolateral route through the dermis, entering the ectoderm through small holes in the basal lamina, or a ventromedial route between the somites and the neural tube. The exception to this is the melanophores of the retinal pigmented epithelium of the eye. These are not derived from the neural crest. Instead, an outpouching of the neural tube generates the optic cup, which, in turn, forms the retina.[citation needed]
When and how multipotent chromatophore precursor cells (called chromatoblasts) develop into their daughter subtypes is an area of ongoing research. It is known in zebrafish embryos, for example, that by 3 days after fertilization each of the cell classes found in the adult fish—melanophores, xanthophores and iridophores—are already present. Studies using mutant fish have demonstrated that transcription factors such as kit, sox10, and mitf are important in controlling chromatophore differentiation.[35] If these proteins are defective, chromatophores may be regionally or entirely absent, resulting in a leucistic disorder.
Practical applications
Chromatophores are sometimes used in applied research. For example, zebrafish larvae are used to study how chromatophores organise and communicate to accurately generate the regular horizontal striped pattern as seen in adult fish.[36] This is seen as a useful model system for understanding patterning in the evolutionary developmental biology field. Chromatophore biology has also been used to model human condition or disease, including melanoma and albinism. Recently, the gene responsible for the melanophore-specific golden zebrafish strain, Slc24a5, was shown to have a human equivalent that strongly correlates with skin colour.[37]
Chromatophores are also used as a biomarker of blindness in cold-blooded species, as animals with certain visual defects fail to background adapt to light environments.[32] Human homologues of receptors that mediate pigment translocation in melanophores are thought to be involved in processes such as appetite suppression and tanning, making them attractive targets for drugs.[26] Therefore, pharmaceutical companies have developed a biological assay for rapidly identifying potential bioactive compounds using melanophores from the African clawed frog.[38] Other scientists have developed techniques for using melanophores as biosensors,[39] and for rapid disease detection (based on the discovery that pertussis toxin blocks pigment aggregation in fish melanophores).[40] Potential military applications of chromatophore-mediated colour changes have been proposed, mainly as a type of active camouflage, which could as in cuttlefish make objects nearly invisible.[41][42]
Cephalopod chromatophores
An infant cuttlefish, using background adaptation to mimic the local environment
Coleoid cephalopods (including octopuses, squids and cuttlefish) have complex multicellular organs that they use to change colour rapidly, producing a wide variety of bright colours and patterns. Each chromatophore unit is composed of a single chromatophore cell and numerous muscle, nerve, glial, and sheath cells.[43] Inside the chromatophore cell, pigment granules are enclosed in an elastic sac, called the cytoelastic sacculus. To change colour the animal distorts the sacculus form or size by muscular contraction, changing its translucency, reflectivity, or opacity. This differs from the mechanism used in fish, amphibians, and reptiles in that the shape of the sacculus is changed, rather than translocating pigment vesicles within the cell. However, a similar effect is achieved.The energy cost of the complete activation of the chromatophore system is very high, being nearly as much as all the energy used by an octopus at rest.[44]
Octopuses and most cuttlefish[45] can operate chromatophores in complex, undulating chromatic displays, resulting in a variety of rapidly changing colour schemata. The nerves that operate the chromatophores are thought to be positioned in the brain in a pattern isomorphic to that of the chromatophores they each control. This means the pattern of colour change functionally matches the pattern of neuronal activation. This may explain why, as the neurons are activated in iterative signal cascade, one may observe waves of colour changing.[46] Like chameleons, cephalopods use physiological colour change for social interaction. They are also among the most skilled at camouflage, having the ability to match both the colour distribution and the texture of their local environment with remarkable accuracy.
↑ Scott M. Boback & Lynn M. Siefferman (2010). "Variation in Color and Color Change in Island and Mainland Boas (Boa constrictor)". Journal of Herpetology. 44 (4): 506–515. doi:10.1670/09-026.1. S2CID53634890.
↑ Aristotle. Historia Animalium. IX, 622a: 2-10. About 400 BC. Cited in Luciana Borrelli, Francesca Gherardi, Graziano Fiorito. A catalogue of body patterning in Cephalopoda. Firenze University Press, 2006. AbstractArchived 2018-02-06 at the Wayback MachineGoogle books
↑ Sangiovanni, G (1819). "Descrizione di un particolare sistema di organi cromoforo espansivo-dermoideo e dei fenomeni che esso produce, scoperto nei molluschi cefaloso". G. Enciclopedico Napoli. 9: 1–13.
↑ Darwin, Charles (1860). "Chapter 1. Habits of a Sea-slug and Cuttle-fish". Journal Of Researches Into The Natural History And Geology Of The Countries Visited During The Voyage Round The World Of H.M.S. 'Beagle' Under The Command Of Captain Fitz Roy, R.N. John Murray, London. p.7.
↑ Fox, DL. Animal Biochromes and Structural Colors: Physical, Chemical, Distributional & Physiological Features of Colored Bodies in the Animal World. University of California Press, Berkeley, 1976. ISBN0-520-02347-1
↑ Bagnara JT. Comparative Anatomy and Physiology of Pigment Cells in Nonmammalian Tissues. In: The Pigmentary System: Physiology and Pathophysiology, Oxford University Press, 1998. ISBN0-19-509861-7
↑ Taylor, JD. (1969). "The effects of intermedin on the ultrastructure of amphibian iridophores". Gen Comp Endocrinol. 12 (3): 405–16. doi:10.1016/0016-6480(69)90157-9. PMID5769930.
↑ Morrison, RL. (1995). "A transmission electron microscopic (TEM) method for determining structural colors reflected by lizard iridophores". Pigment Cell Res. 8 (1): 28–36. doi:10.1111/j.1600-0749.1995.tb00771.x. PMID7792252.
↑ Ito, S; Wakamatsu, K. (2003). "Quantitative analysis of eumelanin and pheomelanin in humans, mice, and other animals: a comparative review". Pigment Cell Res. 16 (5): 523–31. doi:10.1034/j.1600-0749.2003.00072.x. PMID12950732.
↑ Goda M, Ohata M, Ikoma H, Fujiyoshi Y, Sugimoto M, Fujii R (2011). "Integumental reddish-violet coloration owing to novel dichromatic chromatophores in the teleost fish, Pseudochromis diadema". Pigment Cell Melanoma Res. 24 (4): 614–7. doi:10.1111/j.1755-148X.2011.00861.x. PMID21501419. S2CID3299089.
1 2 Logan, DW; Bryson-Richardson, RJ; Pagán, KE; Taylor, MS; Currie, PD; Jackson, IJ (2003). "The structure and evolution of the melanocortin and MCH receptors in fish and mammals". Genomics. 81 (2): 184–91. doi:10.1016/S0888-7543(02)00037-X. PMID12620396.
↑ Valverde, P; Healy, E; Jackson, I; Rees, JL; Thody, AJ (1995). "Variants of the melanocyte-stimulating hormone receptor gene are associated with red hair and fair skin in humans". Nat Genet. 11 (3): 328–30. doi:10.1038/ng1195-328. PMID7581459. S2CID7980311.
↑ Hansford, Dave (August 6, 2008). "Cuttlefish Change Color, Shape-Shift to Elude Predators". National Geographic News. Wellington, New Zealand. Archived from the original on August 10, 2008. [...] cuttlefish have instead relied on invisibility, a talent that may have applications for human technology. Norman said the military has shown interest in cuttlefish camouflage with a view to one day incorporating similar mechanisms in soldiers' uniforms.
↑ Demski, LS (1992). "Chromatophore systems in teleosts and cephalopods: a levels oriented analysis of convergent systems". Brain, Behavior and Evolution. 40 (2–3): 141–56. doi:10.1159/000113909. PMID1422807.
Melanin is a family of biomolecules organized as oligomers or polymers, which among other functions provide the pigments of many organisms. Melanin pigments are produced in a specialized group of cells known as melanocytes.
Melanocytes are melanin-producing neural crest-derived cells located in the bottom layer of the skin's epidermis, the middle layer of the eye, the inner ear, vaginal epithelium, meninges, bones, and heart found in many mammals and birds. Melanin is a dark pigment primarily responsible for skin color. Once synthesized, melanin is contained in special organelles called melanosomes which can be transported to nearby keratinocytes to induce pigmentation. Thus darker skin tones have more melanosomes present than lighter skin tones. Functionally, melanin serves as protection against UV radiation. Melanocytes also have a role in the immune system.
Melanism is the congenital excess of melanin in an organism resulting in dark pigment.
A melanosome is an organelle found in animal cells and is the site for synthesis, storage and transport of melanin, the most common light-absorbing pigment found in the animal kingdom. Melanosomes are responsible for color and photoprotection in animal cells and tissues.
The alpine salamander is a black salamander that can be found in the French Alps, and through the mountainous range in Europe. It is a member of the genus Salamandra. Their species name, atra, may be derived from the Latin ater, meaning dull black. The salamanders' coloration has evolved over time, as some species are completely monochrome black and others have yellow spotting and marks. Their life expectancy is at least 10 years. There are four subspecies of the alpine salamander, with varied distribution and physical coloration. Unlike other salamanders, whose larvae are developed in water, the alpine salamander and its subspecies are a fully terrestrial species in life and gestation. They give birth to live young.
Equine coat color genetics determine a horse's coat color. Many colors are possible, but all variations are produced by changes in only a few genes. Bay is the most common color of horse, followed by black and chestnut. A change at the agouti locus is capable of turning bay to black, while a mutation at the extension locus can turn bay or black to chestnut.
The Pacific tree frog, also known as the Pacific chorus frog, has a range spanning the Pacific Northwest, from Northern California, Oregon, and Washington to British Columbia in Canada and extreme southern Alaska. They live from sea level to more than 10,000 feet in many types of habitats, reproducing in aquatic settings. They occur in shades of greens or browns and can change colors over periods of hours and weeks.
Biological pigments, also known simply as pigments or biochromes, are substances produced by living organisms that have a color resulting from selective color absorption. Biological pigments include plant pigments and flower pigments. Many biological structures, such as skin, eyes, feathers, fur and hair contain pigments such as melanin in specialized cells called chromatophores. In some species, pigments accrue over very long periods during an individual's lifespan.
The common cuttlefish or European common cuttlefish is one of the largest and best-known cuttlefish species. They are a migratory species that spend the summer and spring inshore for spawning and then move to depths of 100–200 metres (330–660 ft) during autumn and winter. They grow to 49 centimetres (19 in) in mantle length and 4 kilograms (8.8 lb) in weight. Animals from subtropical seas are smaller and rarely exceed 30 centimetres (12 in) in mantle length.
The pharaoh cuttlefish is a large cuttlefish species, growing to 42 cm in mantle length and 5 kg in weight.
Animal colouration is the general appearance of an animal resulting from the reflection or emission of light from its surfaces. Some animals are brightly coloured, while others are hard to see. In some species, such as the peafowl, the male has strong patterns, conspicuous colours and is iridescent, while the female is far less visible.
The genetic basis of coat colour in the Labrador Retriever has been found to depend on several distinct genes. The interplay among these genes is used as an example of epistasis.
Neocaridina davidi is a freshwater shrimp originating from eastern China and northern Taiwan and introduced in the rest of Taiwan, Japan, and Hawaii, which is commonly kept in aquaria. The natural coloration of the shrimp is green-brown, though a wide variety of color morphs exist, including red, yellow, orange, green, blue, violet and black shrimp. Full-grown shrimp reach about 4 centimetres (1.6 in) long. N. davidi shrimp are omnivores that may live 1–2 years. These shrimps have previously been classified as Neocaridina heteropoda and Neocaridina denticulata sinensis, however are now known as Neocaridina davidi which is based on the oldest known published description of the species.
Axanthism is a mutation that interferes with an animal's ability to produce yellow pigment. The mutation affects the amount of xanthophores and carotenoid vesicles, sometimes causing them to be completely absent. Erythrophores and iridophores, which are responsible for red coloration and light reflecting pigments respectively, may also be affected. Axanthism is most obvious in green animals, specifically amphibians, making them appear blue. Green coloration in animals is caused by iridiphores reflecting blue wavelengths of light back through the carotenoids in the xanthophores. In the absence of xanthophores and carotenoids, the blue light is unaltered and reflected back normally. Animals that are normally yellow will appear white if affected with axanthism.
Cuttlefish, or cuttles, are marine molluscs of the suborder Sepiina. They belong to the class Cephalopoda which also includes squid, octopuses, and nautiluses. Cuttlefish have a unique internal shell, the cuttlebone, which is used for control of buoyancy.
Amelanism is a pigmentation abnormality characterized by the lack of pigments called melanins, commonly associated with a genetic loss of tyrosinase function. Amelanism can affect fish, amphibians, reptiles, birds, and mammals including humans. The appearance of an amelanistic animal depends on the remaining non-melanin pigments. The opposite of amelanism is melanism, a higher percentage of melanin.
The melanocortin 1 receptor (MC1R), also known as melanocyte-stimulating hormone receptor (MSHR), melanin-activating peptide receptor, or melanotropin receptor, is a G protein–coupled receptor that binds to a class of pituitary peptide hormones known as the melanocortins, which include adrenocorticotropic hormone (ACTH) and the different forms of melanocyte-stimulating hormone (MSH). It is coupled to Gαs and upregulates levels of cAMP by activating adenylyl cyclase in cells expressing this receptor. It is normally expressed in skin and melanocytes, and to a lesser degree in periaqueductal gray matter, astrocytes and leukocytes. In skin cancer, MC1R is highly expressed in melanomas but not carcinomas.
The Atlantic trumpetfish, also referred to as Atlantic coronetfish, is a species of trumpetfish in the family Aulostomidae. It is a tropical marine fish found in shallow coastal waters in the eastern Atlantic Ocean from Mauritania to Namibia. Like other trumpetfish, they eat mainly small fish and often shadow other piscivores while hunting.
Albinism is the congenital absence of melanin in an animal or plant resulting in white hair, feathers, scales and skin and reddish pink or blue eyes. Individuals with the condition are referred to as albinos.
Fish coloration, a subset of animal coloration, is extremely diverse. Fish across all taxa vary greatly in their coloration through special mechanisms, mainly pigment cells called chromatophores. Fish can have any colors of the visual spectrum on their skin, evolutionarily derived for many reasons. There are three factors to coloration, brightness, hue, and saturation. Fish coloration has three proposed functions: thermoregulation, intraspecific communication, and interspecific communication. Fishes' diverse coloration is possibly derivative of the fact that "fish most likely see colors very differently than humans".
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.