In mycology, the terms teleomorph, anamorph, and holomorph apply to portions of the life cycles of fungi in the phyla Ascomycota and Basidiomycota:
Fungi are classified primarily based on the structures associated with sexual reproduction, which tend to be evolutionarily conserved. However, many fungi reproduce only asexually, and cannot easily be classified based on sexual characteristics; some produce both asexual and sexual states. These species are often members of the Ascomycota, but a few of them belong to the Basidiomycota. Even among fungi that reproduce both sexually and asexually, often only one method of reproduction can be observed at a specific point in time or under specific conditions. Additionally, fungi typically grow in mixed colonies and sporulate amongst each other. These facts have made it very difficult to link the various states of the same fungus.
Fungi that are not known to produce a teleomorph were historically placed into an artificial phylum, the "Deuteromycota," also known as "fungi imperfecti," simply for convenience. Some workers hold that this is an obsolete concept, and that molecular phylogeny allows accurate placement of species which are known from only part of their life cycle. Others retain the term "deuteromycetes," but give it a lowercase "d" and no taxonomic rank. [1]
Historically, Article 59 of the International Code of Botanical Nomenclature permitted mycologists to give asexually reproducing fungi (anamorphs) separate names from their sexual states (teleomorphs); [2] but this practice was discontinued as of 1 January 2013. [3]
The dual naming system can be confusing. [4] However, it is essential for workers in plant pathology, mold identification, medical mycology, and food microbiology, fields in which asexually reproducing fungi are commonly encountered.[ clarification needed ]
The separate names for anamorphs of fungi with a pleomorphic life-cycle has been an issue of debate since the phenomenon was recognized in the mid-19th century. [3] This was even before the first international rules for botanical nomenclature were issued in 1867. [3] Special provisions are to be found in the earliest Codes, which were then modified several times, and often substantially. [3] The rules have been updated regularly and become increasingly complex, and by the mid-1970s they were being interpreted in different ways by different mycologists – even ones working on the same genus. [3] Following intensive discussions under the auspices of the International Mycological Association, drastic changes were made at the International Botanical Congress in 1981 to clarify and simplify the procedures – and the new terms anamorph, teleomorph, and holomorph entered general use. [3] An unfortunate effect of the simplification was that many name changes had to be made, including for some well-known and economically important species; at that date, the conservation of species names was not allowed under the Code. [3]
Unforeseen in the 1970s, when the 1981 provisions were crafted, was the impact of molecular systematics. [3] A decade later, it was starting to become obvious that fungi with no known sexual stage could confidently be placed in genera which were typified by species in which the sexual stage was known. [3] This possibility of abandoning the dual nomenclatural system was debated at subsequent International Mycological Congresses and on other occasions, and the need for change was increasingly recognized. [3] [5] At the International Botanical Congress in Vienna in 2005, some minor modifications were made which allowed anamorph-typified names to be epitypified by material showing the sexual stage when it was discovered, and for that anamorph name to continue to be used. [3]
The 1995 edition of the influential Ainsworth and Bisby’s Dictionary of the Fungi sought to replace the term anamorph with mitosporic fungus and teleomorph with meiosporic fungus, based on the idea that the fundamental distinction is whether mitosis or meiosis preceded sporulation. This is a controversial choice because it is not clear that the morphological differences which traditionally define anamorphs and teleomorphs line up completely with sexual practices, or whether those sexual practices are sufficiently well understood in some cases. [1]
The Vienna Congress (2005) established a Special Committee to investigate the issue further, but it was unable to reach a consensus. [3] Matters were becoming increasingly desperate as mycologists using molecular phylogenetic approaches started to ignore the provisions, or interpret them in different ways. [3]
The International Botanical Congress in Melbourne in July 2011 made a change in the International Code of Nomenclature for algae, fungi, and plants and adopted the principle "one fungus, one name". [3] After 1 January 2013, one fungus can only have one name; the system of permitting separate names to be used for anamorphs then ended. [3] This means that all legitimate names proposed for a species, regardless of what stage they are typified by, can serve as the correct name for that species. [3] [6]
All names now compete on an equal footing for priority. [3] In order not to render illegitimate the names that had been introduced in the past for separate morphs, it was agreed that these should not be treated as superfluous alternative names in the sense of the Code. [3] It was further decided that no anamorph-typified name should be taken up to displace a widely used teleomorph-typified name without the case's having been considered by the General Committee established by the Congress. [3] Recognizing that there were cases in some groups of fungi where there could be many names that might merit formal retention or rejection, a new provision [3] was introduced: Lists of names can be submitted to the General Committee and, after due scrutiny, names accepted on those lists are to be treated as conserved over competing synonyms (and listed as Appendices to the Code). [3] Lichen-forming fungi (but not lichenicolous fungi) had always been excluded from the provisions permitting dual nomenclature. [3]
The problem of choosing one name among many remains to be examined for many large, agriculturally or medically-important genera like Aspergillus and Fusarium . Articles have been published on such specific genera to propose ways to define them under the newer rules. [7] [8]

The International Code of Nomenclature for algae, fungi, and plants is the set of rules and recommendations dealing with the formal botanical names that are given to plants, fungi and a few other groups of organisms, all those "traditionally treated as algae, fungi, or plants". It was formerly called the International Code of Botanical Nomenclature (ICBN); the name was changed at the International Botanical Congress in Melbourne in July 2011 as part of the Melbourne Code which replaced the Vienna Code of 2005.
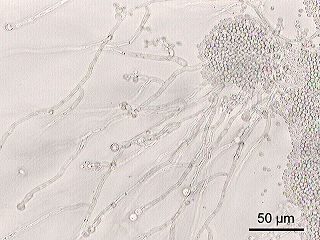
Saccharomycotina is a subdivision (subphylum) of the division (phylum) Ascomycota in the kingdom Fungi. It comprises most of the ascomycete yeasts. The members of Saccharomycotina reproduce by budding and they do not produce ascocarps.
Tolypocladium inflatum is an ascomycete fungus originally isolated from a Norwegian soil sample that, under certain conditions, produces the immunosuppressant drug ciclosporin. In its sexual stage (teleomorph) it is a parasite on scarab beetles. It forms a small, compound ascocarp that arises from the cadaver of its host beetle. In its asexual stage (anamorph) it is a white mold that grows on soil. It is much more commonly found in its asexual stage and this is the stage that was originally given the name Tolypocladium inflatum.
Hyphomycetes are a form classification of fungi, part of what has often been referred to as fungi imperfecti, Deuteromycota, or anamorphic fungi. Hyphomycetes lack closed fruit bodies, and are often referred to as moulds. Most hyphomycetes are now assigned to the Ascomycota, on the basis of genetic connections made by life-cycle studies or by phylogenetic analysis of DNA sequences; many remain unassigned phylogenetically.

Chorioactis is a genus of fungi that contains the single species Chorioactis geaster. The mushroom is commonly known as the devil's cigar or the Texas star in the United States, while in Japan it is called kirinomitake (キリノミタケ). This extremely rare mushroom is notable for its unusual appearance and disjunct distribution; it is found only in select locales in Texas and Japan. The fruit body, which grows on the stumps or dead roots of cedar elms or dead oaks, somewhat resembles a dark brown or black cigar before it splits open radially into a starlike arrangement of four to seven leathery rays. The interior surface of the fruit body bears the spore-bearing tissue known as the hymenium, and is colored white to brown, depending on its age. The fruit body opening can be accompanied by a distinct hissing sound and the release of a smoky cloud of spores.
Conioscyphascus is a fungal genus in the Sordariomycetes class of the Ascomycota. The relationship of this taxon to other taxa within the class is unknown, and it has not yet been placed with certainty into any order or family. The genus, Conioscyphascus contains the single species Conioscyphascus varius, described as new to science in 2004. Until Conioscyphascus gracilis(Munk) Réblová & Seifert was found later in 2004.

Dendrocollybia is a fungal genus in the family Tricholomataceae of the order Agaricales. It is a monotypic genus, containing the single species Dendrocollybia racemosa, commonly known as the branched collybia or the branched shanklet. The somewhat rare species is found in the Northern Hemisphere, including the Pacific Northwest region of western North America, and Europe, where it is included in several Regional Red Lists. It usually grows on the decaying fruit bodies of other agarics—such as Lactarius and Russula—although the host mushrooms may be decayed to the point of being difficult to recognize.

Chlamydosauromyces punctatus is the sole species in the monotypic genus of fungi, Chlamydosauromyces in the family, Onygenaceae. It was found in the skin shed from frilled lizard. This fungus is mesophilic and digests hair. It reproduces both sexually and asexually. The fungus has so far not been reported to be pathogenic.

Sarcoscypha dudleyi, commonly known as the crimson cup or the scarlet cup, is a species of fungus in the family Sarcoscyphaceae of the order Pezizales. In addition to its main distribution in the central to eastern United States, the fungus has also been recorded once in Bulgaria. It has been frequently confused with Sarcoscypha coccinea, but can be distinguished from this and other related species in Sarcoscypha by differences in microscopic characteristics, such as the presence and number of oil droplets in the spores. The species Molliardiomyces dudleyi is an imperfect form of the fungus that lacks a sexually reproductive stage in its life cycle.

Macrotyphula is a genus of clavarioid fungi in the family Phyllotopsidaceae. Basidiocarps are simple, narrowly club-shaped to filiform, sometimes arising from a sclerotium. They typically grow on dead wood or leaf litter, often in swarms.

Ceratobasidium is a genus of fungi in the order Cantharellales. Basidiocarps are effused and the genus is sometimes grouped among the corticioid fungi, though species also retain features of the heterobasidiomycetes. Anamorphic forms were formerly referred to the genus Ceratorhiza, but this is now considered a synonym of Rhizoctonia. Ceratobasidium species, excluding the type, are also now considered synonymous with Rhizoctonia and some species have been transferred to the latter genus. Species are saprotrophic, but several are also facultative plant pathogens, causing a number of commercially important crop diseases. Some are also endomycorrhizal associates of orchids.

Rhizoctonia is a genus of fungi in the order Cantharellales. Species form thin, effused, corticioid basidiocarps, but are most frequently found in their sterile, anamorphic state. Rhizoctonia species are saprotrophic, but some are also facultative plant pathogens, causing commercially important crop diseases. Some are also endomycorrhizal associates of orchids. The genus name was formerly used to accommodate many superficially similar, but unrelated fungi.

Ditangium is a genus of fungi in the family Sebacinaceae. Species form gelatinous basidiocarps (fruitbodies) on wood, often with similarly gelatinous anamorphs. The former were previously referred to the genus Craterocolla, whilst the latter were referred to Ditangium. Following changes to the International Code of Nomenclature for algae, fungi, and plants, the practice of giving different names to teleomorph and anamorph forms of the same fungus was discontinued, meaning that Craterocolla became a synonym of the earlier name Ditangium.
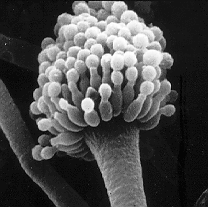
The fungi imperfecti or imperfect fungi are fungi which do not fit into the commonly established taxonomic classifications of fungi that are based on biological species concepts or morphological characteristics of sexual structures because their sexual form of reproduction has never been observed. They are known as imperfect fungi because only their asexual and vegetative phases are known. They have asexual form of reproduction, meaning that these fungi produce their spores asexually, in the process called sporogenesis.

Helicobasidium is a genus of fungi in the subdivision Pucciniomycotina. Basidiocarps are corticioid (patch-forming) and are typically violet to purple. Microscopically they have auricularioid basidia. Asexual anamorphs, formerly referred to the genus Thanatophytum, produce sclerotia. Conidia-bearing anamorphs are parasitic on rust fungi and are currently still referred to the genus Tuberculina.
Blastomyces is a genus of fungi in the order Onygenales. Species are known human pathogens and show thermal dimorphism, converting from hyphal states under saprobic conditions to yeast-like states under pathogenic conditions. They are the causative agents of blastomycosis, a systemic mycosis in immunocompromised patients.
Luella Kayla Weresub, Ph.D. was a world authority on the botanical nomenclature of fungi, especially corticioid fungi and sclerotium-producing basidiomycetes. She was a mycologist at the Central Experimental Farm, Ottawa, with Canada’s federal department of agriculture. Her influence on Canadian mycology and her concern with public education are recognized in the Canadian Botanical Association’s annual Luella K. Weresub lecture in Mycology and the Weresub Prize awarded for the best student paper published by a Canadian student in mycology.
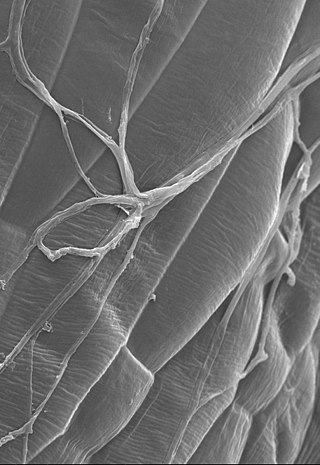
Epichloë festucae is a systemic and seed-transmissible endophytic fungus of cool season grasses.

Botryobasidium is a genus of corticioid fungi belonging to the order Cantharellales. Basidiocarps are ephemeral and typically form thin, web-like, white to cream, effused patches on the underside of fallen branches, logs, and leaf litter. Several species form anamorphs producing chlamydospores. All species are wood- or litter-rotting saprotrophs and the genus has a worldwide distribution.
Conioscypha is a genus of terrestrial and freshwater fungi in the monotypic family Conioscyphaceae and the monotypic order Conioscyphales. They are found on decayed wood, leaves, or bamboo stems. Except for Conioscypha japonica which was isolated from dog skin fragments and hair in 2017.
This article incorporates CC-BY-3.0 text from the reference [3]
| Branch | |||||
|---|---|---|---|---|---|
| Structure |
| ||||
| Growth patterns | |||||
| List | |||||