Evolution is the change in the heritable characteristics of biological populations over successive generations. It occurs when evolutionary processes such as natural selection and genetic drift act on genetic variation, resulting in certain characteristics becoming more or less common within a population over successive generations. The process of evolution has given rise to biodiversity at every level of biological organisation.

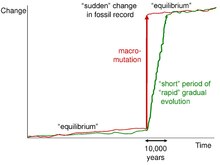
In evolutionary biology, punctuated equilibrium is a theory that proposes that once a species appears in the fossil record, the population will become stable, showing little evolutionary change for most of its geological history. This state of little or no morphological change is called stasis. When significant evolutionary change occurs, the theory proposes that it is generally restricted to rare and geologically rapid events of branching speciation called cladogenesis. Cladogenesis is the process by which a species splits into two distinct species, rather than one species gradually transforming into another.
Speciation is the evolutionary process by which populations evolve to become distinct species. The biologist Orator F. Cook coined the term in 1906 for cladogenesis, the splitting of lineages, as opposed to anagenesis, phyletic evolution within lineages. Charles Darwin was the first to describe the role of natural selection in speciation in his 1859 book On the Origin of Species. He also identified sexual selection as a likely mechanism, but found it problematic.

Theoretical ecology is the scientific discipline devoted to the study of ecological systems using theoretical methods such as simple conceptual models, mathematical models, computational simulations, and advanced data analysis. Effective models improve understanding of the natural world by revealing how the dynamics of species populations are often based on fundamental biological conditions and processes. Further, the field aims to unify a diverse range of empirical observations by assuming that common, mechanistic processes generate observable phenomena across species and ecological environments. Based on biologically realistic assumptions, theoretical ecologists are able to uncover novel, non-intuitive insights about natural processes. Theoretical results are often verified by empirical and observational studies, revealing the power of theoretical methods in both predicting and understanding the noisy, diverse biological world.

Evolutionary biology is the subfield of biology that studies the evolutionary processes that produced the diversity of life on Earth. It is also defined as the study of the history of life forms on Earth. Evolution holds that all species are related and gradually change over generations. In a population, the genetic variations affect the phenotypes of an organism. These changes in the phenotypes will be an advantage to some organisms, which will then be passed on to their offspring. Some examples of evolution in species over many generations are the peppered moth and flightless birds. In the 1930s, the discipline of evolutionary biology emerged through what Julian Huxley called the modern synthesis of understanding, from previously unrelated fields of biological research, such as genetics and ecology, systematics, and paleontology.

Cladogenesis is an evolutionary splitting of a parent species into two distinct species, forming a clade.

This is a list of topics in evolutionary biology.
In biology, adaptation has three related meanings. Firstly, it is the dynamic evolutionary process of natural selection that fits organisms to their environment, enhancing their evolutionary fitness. Secondly, it is a state reached by the population during that process. Thirdly, it is a phenotypic trait or adaptive trait, with a functional role in each individual organism, that is maintained and has evolved through natural selection.

Genetics and the Origin of Species is a 1937 book by the Ukrainian-American evolutionary biologist Theodosius Dobzhansky. It is regarded as one of the most important works of modern synthesis and was one of the earliest. The book popularized the work of population genetics to other biologists and influenced their appreciation for the genetic basis of evolution. In his book, Dobzhansky applied the theoretical work of Sewall Wright (1889–1988) to the study of natural populations, allowing him to address evolutionary problems in a novel way during his time. Dobzhansky implements theories of mutation, natural selection, and speciation throughout his book to explain the habits of populations and the resulting effects on their genetic behavior. The book explains evolution in depth as a process over time that accounts for the diversity of all life on Earth. The study of evolution was present, but greatly neglected at the time. Dobzhansky illustrates that evolution regarding the origin and nature of species during this time in history was deemed mysterious, but had expanding potential for progress to be made in its field.
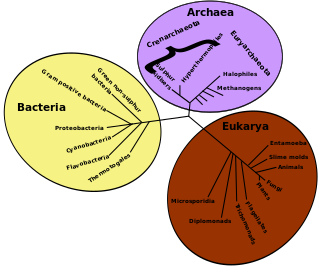
Evolutionary ecology lies at the intersection of ecology and evolutionary biology. It approaches the study of ecology in a way that explicitly considers the evolutionary histories of species and the interactions between them. Conversely, it can be seen as an approach to the study of evolution that incorporates an understanding of the interactions between the species under consideration. The main subfields of evolutionary ecology are life history evolution, sociobiology, the evolution of interspecific interactions and the evolution of biodiversity and of ecological communities.
Niles Eldredge is an American biologist and paleontologist, who, along with Stephen Jay Gould, proposed the theory of punctuated equilibrium in 1972.
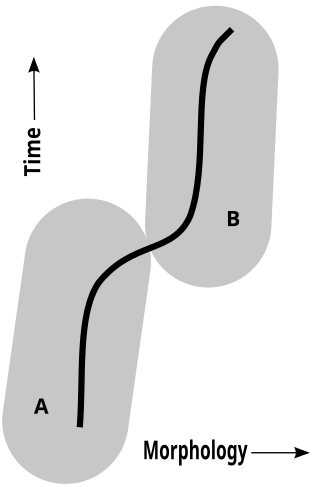
Pseudoextinction of a species occurs when all members of the species are extinct, but members of a daughter species remain alive. The term pseudoextinction refers to the evolution of a species into a new form, with the resultant disappearance of the ancestral form. Pseudoextinction results in the relationship between ancestor and descendant still existing even though the ancestor species no longer exists.
The Red Queen's hypothesis is a hypothesis in evolutionary biology proposed in 1973, that species must constantly adapt, evolve, and proliferate in order to survive while pitted against ever-evolving opposing species. The hypothesis was intended to explain the constant (age-independent) extinction probability as observed in the paleontological record caused by co-evolution between competing species; however, it has also been suggested that the Red Queen hypothesis explains the advantage of sexual reproduction at the level of individuals, and the positive correlation between speciation and extinction rates in most higher taxa.

Dawkins vs. Gould: Survival of the Fittest is a book about the differing views of biologists Richard Dawkins and Stephen Jay Gould by philosopher of biology Kim Sterelny. When published in 2001 it became an international best-seller. A new edition was published in 2007 to include Gould's The Structure of Evolutionary Theory finished shortly before his death in 2002, and recent works by Dawkins. The synopsis below is from the 2007 publication.

In biology, evolution is the process of change in all forms of life over generations, and evolutionary biology is the study of how evolution occurs. Biological populations evolve through genetic changes that correspond to changes in the organisms' observable traits. Genetic changes include mutations, which are caused by damage or replication errors in organisms' DNA. As the genetic variation of a population drifts randomly over generations, natural selection gradually leads traits to become more or less common based on the relative reproductive success of organisms with those traits.
The court jester hypothesis is used in reference to the idea that abiotic forces, rather than biotic competition between species, function as a major driving force behind the processes in evolution which produce speciation. In evolutionary theory, the court-jester hypothesis contrasts the Red Queen hypothesis.

The following outline is provided as an overview of and topical guide to evolution:
The Extended Evolutionary Synthesis (EES) consists of a set of theoretical concepts argued to be more comprehensive than the earlier modern synthesis of evolutionary biology that took place between 1918 and 1942. The extended evolutionary synthesis was called for in the 1950s by C. H. Waddington, argued for on the basis of punctuated equilibrium by Stephen Jay Gould and Niles Eldredge in the 1980s, and was reconceptualized in 2007 by Massimo Pigliucci and Gerd B. Müller.

Frozen Evolution is a 2008 book written by parasitologist Jaroslav Flegr, which aims to explain modern developments in evolutionary biology. It also contains information boxes which clarify important topics in science like peer review, scientific journals, citation metrics, philosophy of science, paradigm shifts, and Occam's razor. Flegr's previous research in toxoplasmosis is also mentioned.

Bruce Smith Lieberman is an American paleontologist.