Transcription is the process of copying a segment of DNA into RNA. The segments of DNA transcribed into RNA molecules that can encode proteins produce messenger RNA (mRNA). Other segments of DNA are transcribed into RNA molecules called non-coding RNAs (ncRNAs).
In molecular biology, the TATA box is a sequence of DNA found in the core promoter region of genes in archaea and eukaryotes. The bacterial homolog of the TATA box is called the Pribnow box which has a shorter consensus sequence.

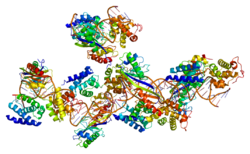
The preinitiation complex is a complex of approximately 100 proteins that is necessary for the transcription of protein-coding genes in eukaryotes and archaea. The preinitiation complex positions RNA polymerase II at gene transcription start sites, denatures the DNA, and positions the DNA in the RNA polymerase II active site for transcription.

General transcription factors (GTFs), also known as basal transcriptional factors, are a class of protein transcription factors that bind to specific sites (promoter) on DNA to activate transcription of genetic information from DNA to messenger RNA. GTFs, RNA polymerase, and the mediator constitute the basic transcriptional apparatus that first bind to the promoter, then start transcription. GTFs are also intimately involved in the process of gene regulation, and most are required for life.

The TATA-binding protein (TBP) is a general transcription factor that binds to a DNA sequence called the TATA box. This DNA sequence is found about 30 base pairs upstream of the transcription start site in some eukaryotic gene promoters.
Transcription factor II D (TFIID) is one of several general transcription factors that make up the RNA polymerase II preinitiation complex. RNA polymerase II holoenzyme is a form of eukaryotic RNA polymerase II that is recruited to the promoters of protein-coding genes in living cells. It consists of RNA polymerase II, a subset of general transcription factors, and regulatory proteins known as SRB proteins. Before the start of transcription, the transcription Factor II D (TFIID) complex binds to the core promoter DNA of the gene through specific recognition of promoter sequence motifs, including the TATA box, Initiator, Downstream Promoter, Motif Ten, or Downstream Regulatory elements.
Eukaryotic transcription is the elaborate process that eukaryotic cells use to copy genetic information stored in DNA into units of transportable complementary RNA replica. Gene transcription occurs in both eukaryotic and prokaryotic cells. Unlike prokaryotic RNA polymerase that initiates the transcription of all different types of RNA, RNA polymerase in eukaryotes comes in three variations, each translating a different type of gene. A eukaryotic cell has a nucleus that separates the processes of transcription and translation. Eukaryotic transcription occurs within the nucleus where DNA is packaged into nucleosomes and higher order chromatin structures. The complexity of the eukaryotic genome necessitates a great variety and complexity of gene expression control.
Transcription factor TFIIA is a nuclear protein involved in the RNA polymerase II-dependent transcription of DNA. TFIIA is one of several general (basal) transcription factors (GTFs) that are required for all transcription events that use RNA polymerase II. Other GTFs include TFIID, a complex composed of the TATA binding protein TBP and TBP-associated factors (TAFs), as well as the factors TFIIB, TFIIE, TFIIF, and TFIIH. Together, these factors are responsible for promoter recognition and the formation of a transcription preinitiation complex (PIC) capable of initiating RNA synthesis from a DNA template.

Transcription initiation factor TFIID subunit 6 is a protein that in humans is encoded by the TAF6 gene.

Transcription initiation factor TFIID subunit 1, also known as transcription initiation factor TFIID 250 kDa subunit (TAFII-250) or TBP-associated factor 250 kDa (p250), is a protein that in humans is encoded by the TAF1 gene.

TAF9 RNA polymerase II, TATA box binding protein (TBP)-associated factor, 32kDa, also known as TAF9, is a protein that in humans is encoded by the TAF9 gene.

Transcription initiation factor TFIID subunit 4 is a protein that in humans is encoded by the TAF4 gene.

Transcription initiation factor TFIID subunit 5 is a protein that in humans is encoded by the TAF5 gene.

Transcription initiation factor TFIID subunit 11 also known as TAFII28, is a protein that in humans is encoded by the TAF11 gene.

Dr1-associated corepressor is a protein that in humans is encoded by the DRAP1 gene.

Transcription factor IIIB 50 kDa subunit (TFIIIB50) also known as b-related factor 2 (BRF-2) is a protein that in humans is encoded by the BRF2 gene.
RNA polymerase II holoenzyme is a form of eukaryotic RNA polymerase II that is recruited to the promoters of protein-coding genes in living cells. It consists of RNA polymerase II, a subset of general transcription factors, and regulatory proteins known as SRB proteins.

The B recognition element (BRE) is a DNA sequence found in the promoter region of most genes in eukaryotes and Archaea. The BRE is a cis-regulatory element that is found immediately near TATA box, and consists of 7 nucleotides. There are two sets of BREs: one (BREu) found immediately upstream of the TATA box, with the consensus SSRCGCC; the other (BREd) found around 7 nucleotides downstream, with the consensus RTDKKKK.
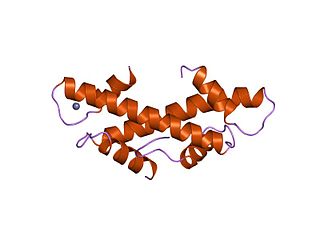
The TBP-associated factors (TAF) are proteins that associate with the TATA-binding protein in transcription initiation. It is a part of the transcription initiation factor TFIID multimeric protein complex. It also makes up many other factors, including SL1. They mediate the formation of the transcription preinitiation complex, a step preceding transcription of DNA to RNA by RNA polymerase II.

Archaeal transcription factor B is a protein family of extrinsic transcription factors that guide the initiation of RNA transcription in organisms that fall under the domain of Archaea. It is homologous to eukaryotic TFIIB and, more distantly, to bacterial sigma factor. Like these proteins, it is involved in forming transcription preinitiation complexes. Its structure includes several conserved motifs which interact with DNA and other transcription factors, notably the single type of RNA polymerase that performs transcription in Archaea.