Runt-related transcription factor 2 (RUNX2) also known as core-binding factor subunit alpha-1 (CBF-alpha-1) is a protein that in humans is encoded by the RUNX2 gene. RUNX2 is a key transcription factor associated with osteoblast differentiation.

Acute myeloblastic leukemia with maturation (M2) is a subtype of acute myeloid leukemia (AML).

ETV6 protein is a transcription factor that in humans is encoded by the ETV6 gene. The ETV6 protein regulates the development and growth of diverse cell types, particularly those of hematological tissues. However, its gene, ETV6 frequently suffers various mutations that lead to an array of potentially lethal cancers, i.e., ETV6 is a clinically significant proto-oncogene in that it can fuse with other genes to drive the development and/or progression of certain cancers. However, ETV6 is also an anti-oncogene or tumor suppressor gene in that mutations in it that encode for a truncated and therefore inactive protein are also associated with certain types of cancers.

Runt-related transcription factor 3 is a protein that in humans is encoded by the RUNX3 gene.

Homeobox protein Hox-A9 is a protein that in humans is encoded by the HOXA9 gene.

GATA2 or GATA-binding factor 2 is a transcription factor, i.e. a nuclear protein which regulates the expression of genes. It regulates many genes that are critical for the embryonic development, self-renewal, maintenance, and functionality of blood-forming, lympathic system-forming, and other tissue-forming stem cells. GATA2 is encoded by the GATA2 gene, a gene which often suffers germline and somatic mutations which lead to a wide range of familial and sporadic diseases, respectively. The gene and its product are targets for the treatment of these diseases.

Core-binding factor subunit beta is a protein that in humans is encoded by the CBFB gene.

CCAAT/enhancer-binding protein alpha is a protein encoded by the CEBPA gene in humans. CCAAT/enhancer-binding protein alpha is a transcription factor involved in the differentiation of certain blood cells. For details on the CCAAT structural motif in gene enhancers and on CCAAT/Enhancer Binding Proteins see the specific page.

Protein CBFA2T1 is a protein that in humans is encoded by the RUNX1T1 gene.

Homeobox protein Hox-A5 is a protein that in humans is encoded by the HOXA5 gene.

MDS1 and EVI1 complex locus protein EVI1 (MECOM) also known as ecotropic virus integration site 1 protein homolog (EVI-1) or positive regulatory domain zinc finger protein 3 (PRDM3) is a protein that in humans is encoded by the MECOM gene. EVI1 was first identified as a common retroviral integration site in AKXD murine myeloid tumors. It has since been identified in a plethora of other organisms, and seems to play a relatively conserved developmental role in embryogenesis. EVI1 is a nuclear transcription factor involved in many signaling pathways for both coexpression and coactivation of cell cycle genes.

Sal-like protein 4(SALL4) is a transcription factor encoded by a member of the Spalt-like (SALL) gene family, SALL4. The SALL genes were identified based on their sequence homology to Spalt, which is a homeotic gene originally cloned in Drosophila melanogaster that is important for terminal trunk structure formation in embryogenesis and imaginal disc development in the larval stages. There are four human SALL proteins with structural homology and playing diverse roles in embryonic development, kidney function, and cancer. The SALL4 gene encodes at least three isoforms, termed A, B, and C, through alternative splicing, with the A and B forms being the most studied. SALL4 can alter gene expression changes through its interaction with many co-factors and epigenetic complexes. It is also known as a key embryonic stem cell (ESC) factor.

K(lysine) acetyltransferase 6A (KAT6A), is an enzyme that, in humans, is encoded by the KAT6A gene. This gene is located on human chromosome 8, band 8p11.21.
Protein CBFA2T2 is a protein that in humans is encoded by the CBFA2T2 gene.

Protein CBFA2T3 is a protein that in humans is encoded by the CBFA2T3 gene.

ETS-related transcription factor Elf-4 is a protein that in humans is encoded by the ELF4 gene.
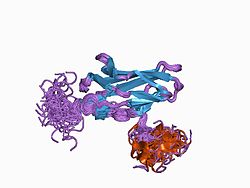
The Runt domain is an evolutionary conserved protein domain. The AML1/RUNX1 gene is rearranged by the t(8;21) translocation in acute myeloid leukemia. The gene is highly similar to the Drosophila melanogaster segmentation gene runt and to the mouse transcription factor PEBP2 alpha subunit gene. The region of shared similarity, known as the Runt domain, is responsible for DNA-binding and protein-protein interaction.

Tet methylcytosine dioxygenase 2 (TET2) is a human gene. It resides at chromosome 4q24, in a region showing recurrent microdeletions and copy-neutral loss of heterozygosity (CN-LOH) in patients with diverse myeloid malignancies.
AI-10-49 is a small molecule inhibitor of leukemic oncoprotein CBFβ-SMHHC developed by the laboratory of John Bushweller with efficacy demonstrated by the laboratories of Lucio H. Castilla and Monica Guzman. AI-10-49 allosterically binds to CBFβ-SMMHC and disrupts protein-protein interaction between CBFβ-SMMHC and tumor suppressor RUNX1. This inhibitor is under development as an anti-leukemic drug.

Musashi-2, also known as Musashi RNA binding protein 2, is a protein that in humans is encoded by the MSI2 gene. Like its homologue musashi-1 (MSI1), it is an RNA-binding protein involved in stemness.