Whether a character state is derived or ancestral is called character polarity. Since genealogical classifications are based on synapomorphies, there must be a way to determine which character state is derived and which is ancestral (or what is special and what is general, to use less evolutionarily freighted terminology) without reference to genealogical classifications, to avoid a circular argument. Some features have been recognized as unique to particular taxa for thousands of years (e.g., feathers for birds, or an internal bony skeleton for vertebrates), and these sorts of common-sense presence/absence characters provide a scaffold upon which the polarity of other characters can be inferred: feathered animals form a natural group; things that lack feathers are just the complement - everything else (mammals, sharks, plants, bacteria). Once a taxon called "birds" is recognized that is defined by the synapomorphy "presence of feathers", then the polarity of other characters present at greater or lesser levels of inclusiveness can be discovered and evaluated. This may identify larger clades, such as the diapsid skull that defines diapsids, or less inclusive clades, such as the syrinx that defines songbirds.
Examples
Lampreys and sharks share some features, like a nervous system, that are not synapomorphic because they are also shared by invertebrates. In contrast, the presence of jaws and paired appendages[11] in both sharks and dogs, but not in lampreys or close invertebrate relatives, identifies these traits as synapomorphies. This supports the hypothesis that dogs and sharks are more closely related to each other than to lampreys.
Clade analysis
The concept of synapomorphy depends on a given clade in the tree of life. Cladograms are diagrams that depict evolutionary relationships within groups of taxa. These illustrations are accurate predictive device in modern genetics. They are usually depicted in either tree or ladder form. Synapomorphies then create evidence for historical relationships and their associated hierarchical structure. Evolutionarily, a synapomorphy is the marker for the most recent common ancestor of the monophyletic group consisting of a set of taxa in a cladogram.[12] What counts as a synapomorphy for one clade may well be a primitive character or plesiomorphy at a less inclusive or nested clade. For example, the presence of mammary glands is a synapomorphy for mammals in relation to tetrapods but is a symplesiomorphy for mammals in relation to one another—rodents and primates, for example. So the concept can be understood as well in terms of "a character newer than" (autapomorphy) and "a character older than" (plesiomorphy) the apomorphy: mammary glands are evolutionarily newer than vertebral column, so mammary glands are an autapomorphy if vertebral column is an apomorphy, but if mammary glands are the apomorphy being considered then vertebral column is a plesiomorphy.[citation needed]
Relations to other terms
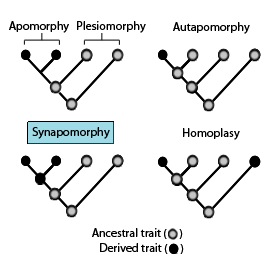
These phylogenetic terms are used to describe different patterns of ancestral and derived character ortraitstates as stated in the above diagram in association with apomorphies and synapomorphies.[13][14]
Symplesiomorphy – an ancestral trait shared by two or more taxa.
Plesiomorphy – a symplesiomorphy discussed in reference to a more derived state.
Pseudoplesiomorphy – a trait that cannot be identified as either a plesiomorphy or an apomorphy that is a reversal.[15]
Reversal – a loss of derived trait present in ancestor and the reestablishment of a plesiomorphic trait.
Convergence – independent evolution of a similar trait in two or more taxa.
Apomorphy – a derived trait. Apomorphy shared by two or more taxa and inherited from a common ancestor is synapomorphy. Apomorphy unique to a given taxon is autapomorphy.[16][17][18][19]
Synapomorphy/homology – a derived trait that is found in some or all terminal groups of a clade, and inherited from a common ancestor, for which it was an autapomorphy (i.e., not present in its immediate ancestor).
Underlying synapomorphy – a synapomorphy that has been lost again in many members of the clade. If lost in all but one, it can be hard to distinguish from an autapomorphy.
Autapomorphy – a distinctive derived trait that is unique to a given taxon or group.[20]
Homoplasy in biological systematics is when a trait has been gained or lost independently in separate lineages during evolution. This convergent evolution leads to species independently sharing a trait that is different from the trait inferred to have been present in their common ancestor.[21][22][23]
Reverse homoplasy – trait present in an ancestor but not in direct descendants that reappears in later descendants.[25]
Hemiplasy is the case where a character that appears homoplastic given the species tree actually has a single origin on the associated gene tree.[26][27] Hemiplasy reflects gene tree-species tree discordance due to the multispecies coalescent.
↑ Futuyma, Douglas J.; Kirkpatrick, Mark (2017). "Tree of life". Evolution (4thed.). Sunderland, Mass.: Sinauer Associates. pp.27–53.
1 2 Futuyma, Douglas J.; Kirkpatrick, Mark (2017). "Phylogeny: The unity and diversity of life". Evolution (4thed.). Sunderland, Mass.: Sinauer Associates. pp.401–429.
↑ "Reconstructing trees: Cladistics". Understanding Evolution. University of California Museum of Paleontology. 5 May 2021. Retrieved 16 October 2021.
↑ Hillis, David M.; Sadava, David; Hill, Richard W.; Price, Mary V. (2014). "Reconstructing and using phylogenies". Principles of Life (2nded.). Sunderland, Mass.: Sinauer Associates. pp.325–342. ISBN978-1464175121.
↑ Novick, Laura R.; Catley, Kefyn M. (December 2007). "Understanding phylogenies in biology: the influence of a Gestalt Perceptual Principle". Journal of Experimental Psychology: Applied. 13 (4): 197–223. doi:10.1037/1076-898X.13.4.197. ISSN1076-898X. PMID18194047.
↑ Appel, Ron D.; Feytmans, Ernest.Bioinformatics: a Swiss Perspective."Chapter 3: Introduction of Phylogenetics and its Molecular Aspects." World Scientific Publishing Company, 1st edition. 2009.
↑ Archie JW (September 1989). "Homoplasy Excess Ratios: New Indices for Measuring Levels of Homoplasy in Phylogenetic Systematics and a Critique of the Consistency Index". Systematic Biology. 38 (1): 253–269. doi:10.2307/2992286. JSTOR2992286.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.