Related Research Articles

A gill is a respiratory organ that many aquatic organisms use to extract dissolved oxygen from water and to excrete carbon dioxide. The gills of some species, such as hermit crabs, have adapted to allow respiration on land provided they are kept moist. The microscopic structure of a gill presents a large surface area to the external environment. Branchia is the zoologists' name for gills.

The respiratory system is a biological system consisting of specific organs and structures used for gas exchange in animals and plants. The anatomy and physiology that make this happen varies greatly, depending on the size of the organism, the environment in which it lives and its evolutionary history. In land animals the respiratory surface is internalized as linings of the lungs. Gas exchange in the lungs occurs in millions of small air sacs; in mammals and reptiles these are called alveoli, and in birds they are known as atria. These microscopic air sacs have a very rich blood supply, thus bringing the air into close contact with the blood. These air sacs communicate with the external environment via a system of airways, or hollow tubes, of which the largest is the trachea, which branches in the middle of the chest into the two main bronchi. These enter the lungs where they branch into progressively narrower secondary and tertiary bronchi that branch into numerous smaller tubes, the bronchioles. In birds the bronchioles are termed parabronchi. It is the bronchioles, or parabronchi that generally open into the microscopic alveoli in mammals and atria in birds. Air has to be pumped from the environment into the alveoli or atria by the process of breathing which involves the muscles of respiration.

The trachea, also known as the windpipe, is a cartilaginous tube that connects the larynx to the bronchi of the lungs, allowing the passage of air, and so is present in almost all air-breathing animals with lungs. The trachea extends from the larynx and branches into the two primary bronchi. At the top of the trachea the cricoid cartilage attaches it to the larynx. The trachea is formed by a number of horseshoe-shaped rings, joined together vertically by overlying ligaments, and by the trachealis muscle at their ends. The epiglottis closes the opening to the larynx during swallowing.
Respiratory failure results from inadequate gas exchange by the respiratory system, meaning that the arterial oxygen, carbon dioxide, or both cannot be kept at normal levels. A drop in the oxygen carried in the blood is known as hypoxemia; a rise in arterial carbon dioxide levels is called hypercapnia. Respiratory failure is classified as either Type 1 or Type 2, based on whether there is a high carbon dioxide level, and can be acute or chronic. In clinical trials, the definition of respiratory failure usually includes increased respiratory rate, abnormal blood gases, and evidence of increased work of breathing. Respiratory failure causes an altered mental status due to ischemia in the brain.
Mechanical ventilation, assisted ventilation or intermittent mandatory ventilation (IMV), is the medical term for artificial ventilation where mechanical means are used to assist or replace spontaneous breathing. This may involve a machine called a ventilator, or the breathing may be assisted manually by a suitably qualified professional, such as an anesthesiologist, Respiratory Therapist, Registered Nurse, paramedic or other first responder, by compressing a bag valve mask device.
The respiratory tract is the subdivision of the respiratory system involved with the process of respiration in mammals. The respiratory tract is lined with respiratory mucosa or respiratory epithelium.

Aquatic respiration is the process whereby an aquatic organism exchanges respiratory gases with water, obtaining oxygen from oxygen dissolved in water and excreting carbon dioxide and some other metabolic waste products into the water.
Gas exchange is the physical process by which gases move passively by diffusion across a surface. For example, this surface might be the air/water interface of a water body, the surface of a gas bubble in a liquid, a gas-permeable membrane, or a biological membrane that forms the boundary between an organism and its extracellular environment.
Exhalation is the flow of the breath out of an organism. In animals, it is the movement of air from the lungs out of the airways, to the external environment during breathing.
In physiology, respiration is the movement of oxygen from the outside environment to the cells within tissues, and the removal of carbon dioxide in the opposite direction.

Hypercapnia (from the Greek hyper = "above" or "too much" and kapnos = "smoke"), also known as hypercarbia and CO2 retention, is a condition of abnormally elevated carbon dioxide (CO2) levels in the blood. Carbon dioxide is a gaseous product of the body's metabolism and is normally expelled through the lungs. Carbon dioxide may accumulate in any condition that causes hypoventilation, a reduction of alveolar ventilation (the clearance of air from the small sacs of the lung where gas exchange takes place) as well as resulting from inhalation of CO2. Inability of the lungs to clear carbon dioxide, or inhalation of elevated levels of CO2, leads to respiratory acidosis. Eventually the body compensates for the raised acidity by retaining alkali in the kidneys, a process known as "metabolic compensation".
The control of ventilation refers to the physiological mechanisms involved in the control of breathing, which is the movement of air into and out of the lungs. Ventilation facilitates respiration. Respiration refers to the utilization of oxygen and balancing of carbon dioxide by the body as a whole, or by individual cells in cellular respiration.

Respiratory arrest is caused by apnea or respiratory dysfunction severe enough it will not sustain the body. Prolonged apnea refers to a patient who has stopped breathing for a long period of time. If the heart muscle contraction is intact, the condition is known as respiratory arrest. An abrupt stop of pulmonary gas exchange lasting for more than five minutes may damage vital organs especially the brain, possibly permanently. Lack of oxygen to the brain causes loss of consciousness. Brain injury is likely if respiratory arrest goes untreated for more than three minutes, and death is almost certain if more than five minutes.
High-frequency ventilation is a type of mechanical ventilation which utilizes a respiratory rate greater than four times the normal value. and very small tidal volumes. High frequency ventilation is thought to reduce ventilator-associated lung injury (VALI), especially in the context of ARDS and acute lung injury. This is commonly referred to as lung protective ventilation. There are different types of high-frequency ventilation. Each type has its own unique advantages and disadvantages. The types of HFV are characterized by the delivery system and the type of exhalation phase.
Insect physiology includes the physiology and biochemistry of insect organ systems.

Opiliones are an order of arachnids and share many common characteristics with other arachnids. However, several differences separate harvestmen from other arachnid orders such as spiders. The bodies of opiliones are divided into two tagmata : the abdomen (opisthosoma) and the cephalothorax (prosoma). Unlike spiders, the juncture between the abdomen and cephalothorax is often poorly defined. Harvestmen have chelicerae, pedipalps and four pairs of legs. Most harvestmen have two eyes, although there are eyeless species.

Breathing is the process of moving air out and in the lungs to facilitate gas exchange with the internal environment, mostly to flush out carbon dioxide and bring in oxygen.

Insect morphology is the study and description of the physical form of insects. The terminology used to describe insects is similar to that used for other arthropods due to their shared evolutionary history. Three physical features separate insects from other arthropods: they have a body divided into three regions, have three pairs of legs, and mouthparts located outside of the head capsule. It is this position of the mouthparts which divides them from their closest relatives, the non-insect hexapods, which includes Protura, Diplura, and Collembola.
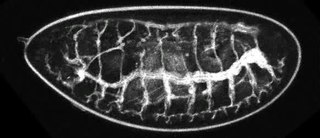
An insect's respiratory system is the system with which it introduces respiratory gases to its interior and performs gas exchange.

A spiracle or stigma is the opening in the exoskeletons of insects and some spiders to allow air to enter the trachea. In the respiratory system of insects, the tracheal tubes primarily deliver oxygen directly into the animals' tissues. The spiracles can be opened and closed in an efficient manner to reduce water loss. This is done by contracting closer muscles surrounding the spiracle. In order to open, the muscle relaxes. The closer muscle is controlled by the central nervous system, but can also react to localized chemical stimuli. Several aquatic insects have similar or alternative closing methods to prevent water from entering the trachea. The timing and duration of spiracle closures can affect the respiratory rates of the organism. Spiracles may also be surrounded by hairs to minimize bulk air movement around the opening, and thus minimize water loss.
References
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 Chown, S.L.; S.W. Nicholson (2004). Insect Physiological Ecology. New York: Oxford University Press. ISBN 0-19-851549-9.
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 Chown SL, Gibbs AG, Hetz SK, Klok CJ, Lighton JR, Marias E (2006). "Discontinuous gas exchange in insects: a clarification of hypotheses and approaches". Physiological and Biochemical Zoology. 79 (2): 333–343. doi:10.1086/499992. PMID 16555192.
- ↑ Nation, J.L. (2002). Insect Physiology and Biochemistry. Boca Raton: C.R.C. Press. ISBN 0-8493-1181-0.
- ↑ Lighton JR, Lovegrove BG (1990). "A temperature-induced switch from diffusive to convective ventilation in the honeybee" (PDF). Journal of Experimental Biology. 154: 509–516.
- 1 2 Chown SL (2002). "Respiratory water loss in insects". Comparative Biochemistry and Physiology A. 133: 791–804. doi:10.1016/s1095-6433(02)00200-3. PMID 12443935.
- ↑ Williams AE, Rose MR, Bradley TJ (1997). "CO2 release patterns in Drosophila melanogaster: the effect of selection for desiccation resistance". Journal of Experimental Biology. 200 (Pt 3): 615–624. PMID 9057311.
- 1 2 Lighton JR, Berrigan D (1995). "Questioning paradigms: caste-specific ventiation in harvester ants, Messor pergandei and M. julianus (Hymenoptera: Formicidae)". Journal of Experimental Biology. 198 (Pt 2): 521–530. PMID 9318205.
- ↑ Lighton JRB (2007). "Respiratory Biology: why insects evolved discontinuous gas exchange". Current Biology. 17 (16): R645–647. doi: 10.1016/j.cub.2007.06.007 . PMID 17714655.
- ↑ Harrison JF, Camazine S, Marden JH, Kirkton SD, Rozo A, Yang X (1995). "Mite not make it home: tracheal mites reduce the safety margin for oxygen delivery of flying honeybees". Journal of Experimental Biology. 204 (Pt 4): 805–13. PMID 11171363.