
An endosymbiont or endobiont is an organism that lives within the body or cells of another organism. Typically the two organisms are in a mutualistic relationship. Examples are nitrogen-fixing bacteria, which live in the root nodules of legumes, single-cell algae inside reef-building corals, and bacterial endosymbionts that provide essential nutrients to insects.

A genome is all the genetic information of an organism. It consists of nucleotide sequences of DNA. The nuclear genome includes protein-coding genes and non-coding genes, other functional regions of the genome such as regulatory sequences, and often a substantial fraction of junk DNA with no evident function. Almost all eukaryotes have mitochondria and a small mitochondrial genome. Algae and plants also contain chloroplasts with a chloroplast genome.
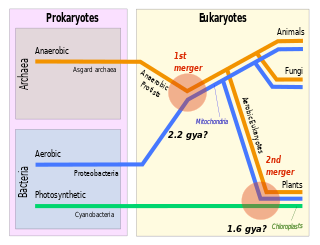
Symbiogenesis is the leading evolutionary theory of the origin of eukaryotic cells from prokaryotic organisms. The theory holds that mitochondria, plastids such as chloroplasts, and possibly other organelles of eukaryotic cells are descended from formerly free-living prokaryotes taken one inside the other in endosymbiosis. Mitochondria appear to be phylogenetically related to Rickettsiales bacteria, while chloroplasts are thought to be related to cyanobacteria.
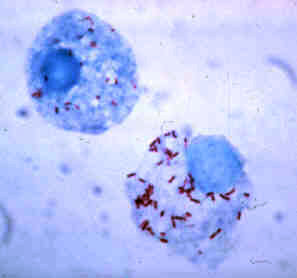
Rickettsia is a genus of nonmotile, gram-negative, nonspore-forming, highly pleomorphic bacteria that may occur in the forms of cocci, bacilli, or threads. The genus was named after Howard Taylor Ricketts in honor of his pioneering work on tick-borne spotted fever.

Horizontal gene transfer (HGT) or lateral gene transfer (LGT) is the movement of genetic material between organisms other than by the ("vertical") transmission of DNA from parent to offspring (reproduction). HGT is an important factor in the evolution of many organisms. HGT is influencing scientific understanding of higher-order evolution while more significantly shifting perspectives on bacterial evolution.
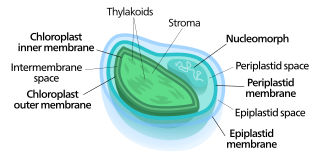
Nucleomorphs are small, vestigial eukaryotic nuclei found between the inner and outer pairs of membranes in certain plastids. They are thought to be vestiges of red and green algal nuclei that were engulfed by a larger eukaryote. Because the nucleomorph lies between two sets of membranes, nucleomorphs support the endosymbiotic theory and are evidence that the plastids containing them are complex plastids. Having two sets of membranes indicate that the plastid, a prokaryote, was engulfed by a eukaryote, an alga, which was then engulfed by another eukaryote, the host cell, making the plastid an example of secondary endosymbiosis.
The Rickettsiales, informally called rickettsias, are an order of small Alphaproteobacteria. They are obligate intracellular parasites, and some are notable pathogens, including Rickettsia, which causes a variety of diseases in humans, and Ehrlichia, which causes diseases in livestock. Another genus of well-known Rickettsiales is the Wolbachia, which infect about two-thirds of all arthropods and nearly all filarial nematodes. Genetic studies support the endosymbiotic theory according to which mitochondria and related organelles developed from members of this group.

Buchnera aphidicola, a member of the Pseudomonadota and the only species in the genus Buchnera, is the primary endosymbiont of aphids, and has been studied in the pea aphid, Acyrthosiphon pisum. Buchnera is believed to have had a free-living, Gram-negative ancestor similar to a modern Enterobacterales, such as Escherichia coli. Buchnera is 3 μm in diameter and has some of the key characteristics of its Enterobacterales relatives, such as a Gram-negative cell wall. However, unlike most other Gram-negative bacteria, Buchnera lacks the genes to produce lipopolysaccharides for its outer membrane. The long association with aphids and the limitation of crossover events due to strictly vertical transmission has seen the deletion of genes required for anaerobic respiration, the synthesis of amino sugars, fatty acids, phospholipids, and complex carbohydrates. This has resulted not only in one of the smallest known genomes of any living organism, but also one of the most genetically stable.

Alphaproteobacteria or α-proteobacteria, also called α-Purple bacteria in earlier literature, is a class of bacteria in the phylum Pseudomonadota. The Magnetococcales and Mariprofundales are considered basal or sister to the Alphaproteobacteria. The Alphaproteobacteria are highly diverse and possess few commonalities, but nevertheless share a common ancestor. Like all Proteobacteria, its members are gram-negative, although some of its intracellular parasitic members lack peptidoglycan and are consequently gram variable.

Paulinella is a genus of at least eleven species including both freshwater and marine amoeboids. Like many members of euglyphids it is covered by rows of siliceous scales, and use filose pseudopods to crawl over the substrate of the benthic zone.

A prokaryote is a single-celled organism whose cell lacks a nucleus and other membrane-bound organelles. The word prokaryote comes from the Ancient Greek πρό (pró), meaning 'before', and κάρυον (káruon), meaning 'nut' or 'kernel'. In the earlier two-empire system arising from the work of Édouard Chatton, prokaryotes were classified within the empire Prokaryota. However, in the three-domain system, based upon molecular phylogenetics, prokaryotes are divided into two domains: Bacteria and Archaea. A third domain, Eukaryota, consists of organisms with nuclei.
The CoRR hypothesis states that the location of genetic information in cytoplasmic organelles permits regulation of its expression by the reduction-oxidation ("redox") state of its gene products.

Acyrthosiphon pisum, commonly known as the pea aphid, is a sap-sucking insect in the family Aphididae. It feeds on several species of legumes worldwide, including forage crops, such as pea, clover, alfalfa, and broad bean, and ranks among the aphid species of major agronomical importance. The pea aphid is a model organism for biological study whose genome has been sequenced and annotated.

Genome evolution is the process by which a genome changes in structure (sequence) or size over time. The study of genome evolution involves multiple fields such as structural analysis of the genome, the study of genomic parasites, gene and ancient genome duplications, polyploidy, and comparative genomics. Genome evolution is a constantly changing and evolving field due to the steadily growing number of sequenced genomes, both prokaryotic and eukaryotic, available to the scientific community and the public at large.
Bacterial genomes are generally smaller and less variant in size among species when compared with genomes of eukaryotes. Bacterial genomes can range in size anywhere from about 130 kbp to over 14 Mbp. A study that included, but was not limited to, 478 bacterial genomes, concluded that as genome size increases, the number of genes increases at a disproportionately slower rate in eukaryotes than in non-eukaryotes. Thus, the proportion of non-coding DNA goes up with genome size more quickly in non-bacteria than in bacteria. This is consistent with the fact that most eukaryotic nuclear DNA is non-gene coding, while the majority of prokaryotic, viral, and organellar genes are coding. Right now, we have genome sequences from 50 different bacterial phyla and 11 different archaeal phyla. Second-generation sequencing has yielded many draft genomes ; third-generation sequencing might eventually yield a complete genome in a few hours. The genome sequences reveal much diversity in bacteria. Analysis of over 2000 Escherichia coli genomes reveals an E. coli core genome of about 3100 gene families and a total of about 89,000 different gene families. Genome sequences show that parasitic bacteria have 500–1200 genes, free-living bacteria have 1500–7500 genes, and archaea have 1500–2700 genes. A striking discovery by Cole et al. described massive amounts of gene decay when comparing Leprosy bacillus to ancestral bacteria. Studies have since shown that several bacteria have smaller genome sizes than their ancestors did. Over the years, researchers have proposed several theories to explain the general trend of bacterial genome decay and the relatively small size of bacterial genomes. Compelling evidence indicates that the apparent degradation of bacterial genomes is owed to a deletional bias.

The minimal genome is a concept which can be defined as the set of genes sufficient for life to exist and propagate under nutrient-rich and stress-free conditions. Alternatively, it may be defined as the gene set supporting life on an axenic cell culture in rich media, and it is thought what makes up the minimal genome will depend on the environmental conditions that the organism inhabits.
Hamiltonella defensa is a species of bacteria. It is maternally or sexually transmitted and lives as an endosymbiont of whiteflies and aphids, meaning that it lives within a host, protecting its host from attack. It does this through bypassing the host's immune responses by protecting its host against parasitoid wasps. However, H. defensa is only defensive if infected by a virus. H. defensa shows a relationship with Photorhabdus species, together with Regiella insecticola. Together with other endosymbionts, it provides aphids protection against parasitoids. It is known to habitate Bemisia tabaci.
Genomic streamlining is a theory in evolutionary biology and microbial ecology that suggests that there is a reproductive benefit to prokaryotes having a smaller genome size with less non-coding DNA and fewer non-essential genes. There is a lot of variation in prokaryotic genome size, with the smallest free-living cell's genome being roughly ten times smaller than the largest prokaryote. Two of the free-living bacterial taxa with the smallest genomes are Prochlorococcus and Pelagibacter ubique, both highly abundant marine bacteria commonly found in oligotrophic regions. Similar reduced genomes have been found in uncultured marine bacteria, suggesting that genomic streamlining is a common feature of bacterioplankton. This theory is typically used with reference to free-living organisms in oligotrophic environments.

A plastid is a membrane-bound organelle found in plants, algae and other eukaryotic organisms that contribute to the production of pigment molecules. Most plastids are photosynthetic, thus leading to color production and energy storage or production. There are many types of plastids in plants alone, but all plastids can be separated based on the number of times they have undergone endosymbiotic events. Currently there are three types of plastids; primary, secondary and tertiary. Endosymbiosis is reputed to have led to the evolution of eukaryotic organisms today, although the timeline is highly debated.
Reductive evolution is the process by which microorganisms remove genes from their genome. It can occur when bacteria found in a free-living state enter a restrictive state or are completely absorbed by another organism becoming intracellular (symbiogenesis). The bacteria will adapt to survive and thrive in the restrictive state by altering and reducing its genome to get rid of the newly redundant pathways that are provided by the host. In an endosymbiont or symbiogenesis relationship where both the guest and host benefit, the host can also undergo reductive evolution to eliminate pathways that are more efficiently provided for by the guest.



