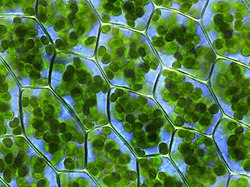
Chloroplasts, proplastids, and differentiation
In land plants, the plastids that contain chlorophyll can perform photosynthesis, thereby creating internal chemical energy from external sunlight energy while capturing carbon from Earth's atmosphere and furnishing the atmosphere with life-giving oxygen. These are the chlorophyll-plastids—and they are named chloroplasts; (see top graphic).
Other plastids can synthesize fatty acids and terpenes, which may be used to produce energy or as raw material to synthesize other molecules. For example, plastid epidermal cells manufacture the components of the tissue system known as plant cuticle, including its epicuticular wax, from palmitic acid —which itself is synthesized in the chloroplasts of the mesophyll tissue. Plastids function to store different components including starches, fats, and proteins. [9]
All plastids are derived from proplastids (also named proplasts [10] ), which are present in the meristematic regions of the plant. Proplastids and young chloroplasts typically divide by binary fission, but more mature chloroplasts also have this capacity.
Plant proplastids (undifferentiated plastids) may differentiate into several forms, depending upon which function they perform in the cell, (see top graphic). They may develop into any of the following variants: [11]
Leucoplasts differentiate into even more specialized plastids, such as:
Depending on their morphology and target function, plastids have the ability to differentiate or redifferentiate between these and other forms.
Plastomes and Chloroplast DNA/ RNA; plastid DNA and plastid nucleoids
Each plastid creates multiple copies of its own unique genome, or plastome, (from 'plastid genome')—which for a chlorophyll plastid (or chloroplast) is equivalent to a 'chloroplast genome', or a 'chloroplast DNA'. [12] [13] The number of genome copies produced per plastid is variable, ranging from 1000 or more in rapidly dividing new cells, encompassing only a few plastids, down to 100 or less in mature cells, encompassing numerous plastids.
A plastome typically contains a genome that encodes transfer ribonucleic acids (tRNA)s and ribosomal ribonucleic acids (rRNAs). It also contains proteins involved in photosynthesis and plastid gene transcription and translation. But these proteins represent only a small fraction of the total protein set-up necessary to build and maintain any particular type of plastid. Nuclear genes (in the cell nucleus of a plant) encode the vast majority of plastid proteins; and the expression of nuclear and plastid genes is co-regulated to coordinate the development and differention of plastids.
Many plastids, particularly those responsible for photosynthesis, possess numerous internal membrane layers. Plastid DNA exists as protein-DNA complexes associated as localized regions within the plastid's inner envelope membrane; and these complexes are called 'plastid nucleoids'. Unlike the nucleus of a eukaryotic cell, a plastid nucleoid is not surrounded by a nuclear membrane. The region of each nucleoid may contain more than 10 copies of the plastid DNA.
Where the proplastid (undifferentiated plastid) contains a single nucleoid region located near the centre of the proplastid, the developing (or differentiating) plastid has many nucleoids localized at the periphery of the plastid and bound to the inner envelope membrane. During the development/ differentiation of proplastids to chloroplasts—and when plastids are differentiating from one type to another—nucleoids change in morphology, size, and location within the organelle. The remodelling of plastid nucleoids is believed to occur by modifications to the abundance of and the composition of nucleoid proteins.
In normal plant cells long thin protuberances called stromules sometimes form—extending from the plastid body into the cell cytosol while interconnecting several plastids. Proteins and smaller molecules can move around and through the stromules. Comparatively, in the laboratory, most cultured cells—which are large compared to normal plant cells—produce very long and abundant stromules that extend to the cell periphery.
In 2014, evidence was found of the possible loss of plastid genome in Rafflesia lagascae, a non-photosynthetic parasitic flowering plant, and in Polytomella , a genus of non-photosynthetic green algae. Extensive searches for plastid genes in both taxons yielded no results, but concluding that their plastomes are entirely missing is still disputed. [14] Some scientists argue that plastid genome loss is unlikely since even these non-photosynthetic plastids contain genes necessary to complete various biosynthetic pathways including heme biosynthesis. [14] [15]
Even with any loss of plastid genome in Rafflesiaceae, the plastids still occur there as "shells" without DNA content, [16] which is reminiscent of hydrogenosomes in various organisms.