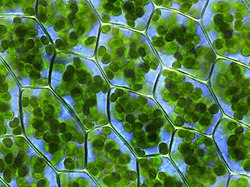
Lichens represent an association between one or more fungalmycobionts and one or more photosynthetic algal or cyanobacterial photobionts. The mycobiont provides protection from predation and desiccation, while the photobiont provides energy in the form of fixed carbon. Cyanobacterial partners are also capable of fixing nitrogen for the fungal partner.[8] Recent work suggests that non-photosynthetic bacterial microbiomes associated with lichens may also have functional significance to lichens.[9]
Most mycobiont partners derive from the ascomycetes, and the largest class of lichenized fungi is Lecanoromycetes.[10] The vast majority of lichens derive photobionts from Chlorophyta (green algae).[8] The co-evolutionary dynamics between mycobionts and photobionts are still unclear, as many photobionts are capable of free-living, and many lichenized fungi display traits adaptive to lichenization such as the capacity to withstand higher levels of reactive oxygen species (ROS), the conversion of sugars to polypols that help withstand dedication, and the downregulation of fungal virulence. However, it is still unclear whether these are derived or ancestral traits.[8]
Currently described photobiont species number about 100, far less than the 19,000 described species of fungal mycobionts, and factors such as geography can predominate over mycobiont preference.[11][12] Phylogenetic analyses in lichenized fungi have suggested that, throughout evolutionary history, there has been repeated loss of photosymbionts, switching of photosymbionts, and independent lichenization events in previously unrelated fungal taxa.[10][13] Loss of lichenization has likely led to the coexistence of non-lichenized fungi and lichenized fungi in lichens.[13]
Sponges
Sponges (phylum Porifera) have a large diversity of photosymbiote associations. Photosymbiosis is found in four classes of Porifera (Demospongiae, Hexactinellida, Homoscleromorpha, and Calcarea), and known photosynthetic partners are cyanobacteria, chloroflexi, dinoflagellates, and red (Rhodophyta) and green (Chlorophyta) algae. Relatively little is known about the evolutionary history of sponge photosymbiois due to a lack of genomic data.[14] However, it has been shown that photosymbiotes are acquired vertically (transmission from parent to offspring) and/or horizontally (acquired from the environment).[15] Photosymbiotes can supply up to half of the host sponge's respiratory demands and can support sponges during times of nutrient stress.[16]
Cnidaria
Members of certain classes in phylum Cnidaria are known for photosymbiotic partnerships. Members of corals (Class Anthozoa) in the orders Hexacorallia and Octocorallia form well-characterized partnerships with the dinoflagellate genus Symbiodinium. Some jellyfish (class Scyphozoa) in the genus Cassiopea (upside-down jellyfish) also possess Symbiodinium. Certain species in the genus Hydra (class Hydrozoa) also harbor green algae and form a stable photosymbiosis.[14]
The evolution of photosymbiosis in corals was likely critical for the global establishment of coral reefs.[17] Corals are likewise adapted to eject damaged photosymbionts that generate high levels of toxic reactive oxygen species, a process known as bleaching.[18] The identity of the Symbiodinium photosymbiont can change in corals, although this depends largely on the mode of transmission: some species vertically transmit their algal partners through their eggs,[19] while other species acquire environmental dinoflagellates as newly-released eggs.[20] Since algae are not preserved in the coral fossil record, understanding the evolutionary history of the symbiosis is difficult.[21]
In freshwater systems, photosymbiosis is present in platyhelminths belonging to the Rhabdocoela group.[14] In this group, members of the Provorticidae, Dalyeliidae, and Typhloplanidae families are symbiotic.[24] Members of Provorticidae likely feed on diatoms and retain their symbionts.[24] Typhloplanidae have symbiotic relationships with the chlorophytes in the genus Chlorella.[33]
Molluscs
Photosymbiosis is taxonomically restricted in Mollusca.[14] Tropical marine bivalves in the Cardiidae family form a symbiotic relationship with the dinoflagellate Symbiodinium.[34] This family boasts large organisms often referred to as giant clams and their large size is attributed to the establishment of these symbiotic relationships. Additionally, the Symbiodinium are hosted extracellularly, which is relatively rare.[35] The only known freshwater bivalve with a symbiotic relationship are in the genus Anodonta which hosts the chlorophyte Chlorella in the gills and mantle of the host.[36] In bivalves, photosymbiosis is thought to have evolved twice, in the genus Anodonta and in the family Cardiidae.[36] However, how it has evolved in Cardiidae could have occurred through different gains or losses in the family.[37]
Gastropods
In gastropods, photosymbiosis can be found in several genera.
The species Strombus gigas hosts Symbiodinium which is acquired during the larval stage, at which point it is a mutualistic relationship.[38] However, during the adult stage, Symbiodinium becomes parasitic as the shell prevents photosynthesis.[39]
Another group of gastropods, heterobranch sea slugs, have two different systems for symbiosis. The first, Nudibranchia, acquire their symbionts through feeding on cnidarian prey that are in symbiotic relationships.[40] In Nudibranchs, photosymbiosis has evolved twice, in Melibe and Aeolidida.[14] In Aeolidida it is likely there have been several gains and losses of photosymbiosis as most genera include both photosymbiotic and non-photosymbiotic species.[14] The second, Sacoglossa, removes chloroplasts from macroalgae when feeding and sequesters them into their digestive tract at which point they are called kleptoplasts.[41] Whether these kleptoplasts maintain their photosynthetic capabilities depends on the host species ability to digest them properly.[42] In this group, functional kleptoplasty has been acquired twice, in Costasiellidae and Plakobranchacea.[43]
Chordates
Photosymbiosis is relatively uncommon in chordate species.[14] One such example of photosymbiosis is in ascidians, the sea squirts. In the genus Didemnidae, 30 species establish symbiotic relationships.[44] The photosynthetic ascidians are associated with cyanobacteria in the genus of Prochloron as well as, in some cases, the species Synechocystis trididemni.[44] The 30 species with a symbiotic relationship span four genera where the congeners (species within the same genus) are primarily non-symbiotic, suggesting multiple origins of photosymbiosis in ascidians.[45]
In addition to sea squirts, embryos of some amphibian species (Ambystoma maculatum, Ambystoma gracile, Ambystoma jeffersonium, Ambystoma trigrinum, Hynobius nigrescens, Lithobates sylvaticus, and Lithobates aurora) form symbiotic relationships with the green alga in the genus of Oophila.[46][47][48] This algae is present in the egg masses of the species, causing them to appear green and providing oxygen and carbohydrates to the embryos.[49] Similarly, little is known about the evolution of symbiosis in amphibians, but there appear to be multiple origins.
Protists
Photosymbiosis has evolved multiple times in the protist taxa Ciliophora, Foraminifera, Radiolaria, Dinoflagellata, and diatoms.[50] Foraminifera and Radiolaria are planktonic taxa that serve as primary producers in open ocean communities.[51] Photosynthetic plankton species associate with the symbiotes of dinoflagellates, diatoms, rhodophytes, chlorophytes, and cyanophytes that can be transferred both vertically and horizontally.[52] In Foraminifera, benthic species will either have a symbiotic relationship with Symbiodinium or retain the chloroplasts present in algal prey species.[50] The planktonic species of Foraminifera associate primarily with Pelagodinium.[50] These species are often considered indicator species due to their bleaching in response to environmental stressors.[53] In the Radiolarian group Acantharia, photosynthetic species inhabit surface waters whereas non-photosynthetic species inhabit deeper waters. Photosynthetic Acantharia are associated with similar microalgae as the Foraminifera groups, but were also found to be associated with Phaeocystis, Heterocapsa, Scrippsiella, and Azadinium which were not previously known to be involved in photosynthetic relationships.[54] In addition, several of the species present in symbiotic relationships with Acantharia were oftentimes identical to the free-living species, suggesting horizontal transfer of symbiotes.[51] This provides insight into the evolutionary patterns responsible for these symbiotic relationships, suggesting that the selection for symbiosis is relatively weak and symbiosis is likely a result of the adaptive capacity of the host plankton species.
↑ Paps, J.; Baguna, J.; Riutort, M. (2009-07-14). "Bilaterian Phylogeny: A Broad Sampling of 13 Nuclear Genes Provides a New Lophotrochozoa Phylogeny and Supports a Paraphyletic Basal Acoelomorpha". Molecular Biology and Evolution. 26 (10): 2397–2406. doi:10.1093/molbev/msp150. ISSN0737-4038. PMID19602542.
↑ SHANNON, THOMAS; ACHATZ, JOHANNES G. (2007-07-12). "Convolutriloba macropyga sp. nov., an uncommonly fecund acoel (Acoelomorpha) discovered in tropical aquaria". Zootaxa. 1525 (1). doi:10.11646/zootaxa.1525.1.1. ISSN1175-5334.
↑ Gschwentner, Robert; Mueller, Johann; Ladurner, Peter; Rieger, Reinhard; Tyler, Seth (2003-02-12). "Unique patterns of longitudinal body-wall musculature in the Acoela (Plathelminthes): the ventral musculature of Convolutriloba longifissura". Zoomorphology. 122 (2): 87–94. doi:10.1007/s00435-003-0074-3. ISSN0720-213X.
↑ Serôdio, João; Silva, Raquel; Ezequiel, João; Calado, Ricardo (2010-07-14). "Photobiology of the symbiotic acoel flatwormSymsagittifera roscoffensis: algal symbiont photoacclimation and host photobehaviour". Journal of the Marine Biological Association of the United Kingdom. 91 (1): 163–171. doi:10.1017/s0025315410001001. ISSN0025-3154.
↑ Taylor, D. L. (May 1971). "On the symbiosis betweenAmphidinium klebsii[Dinophyceae] andAmphiscolops langerhansi[Turbellaria: Acoela]". Journal of the Marine Biological Association of the United Kingdom. 51 (2): 301–313. Bibcode:1971JMBUK..51..301T. doi:10.1017/s0025315400031799. ISSN0025-3154.
↑ Lopes, Rubens Mendes; Silveira, Marina (July 1994). "Symbiosis between a pelagic flatworm and a dinoflagellate from a tropical area: structural observations". Hydrobiologia. 287 (3): 277–284. Bibcode:1994HyBio.287..277L. doi:10.1007/bf00006376. ISSN0018-8158.
↑ Barneah, Orit; Brickner, Itzchak; Hooge, Matthew; Weis, Virginia M.; Benayahu, Yehuda (2007-08-14). "First evidence of maternal transmission of algal endosymbionts at an oocyte stage in a triploblastic host, with observations on reproduction inWaminoa brickneri(Acoelomorpha)". Invertebrate Biology. 126 (2): 113–119. Bibcode:2007InvBi.126..113B. doi:10.1111/j.1744-7410.2007.00082.x. ISSN1077-8306.
↑ Hikosaka-Katayama, Tomoe; Koike, Kanae; Yamashita, Hiroshi; Hikosaka, Akira; Koike, Kazuhiko (September 2012). "Mechanisms of Maternal Inheritance of Dinoflagellate Symbionts in the Acoelomorph WormWaminoa litus". Zoological Science. 29 (9): 559–567. doi:10.2108/zsj.29.559. ISSN0289-0003. PMID22943779.
1 2 PARDY, R. L. (June 1980). "SYMBIOTIC ALGAE AND14C INCORPORATION IN THE FRESHWATER CLAM,ANODONTA". The Biological Bulletin. 158 (3): 349–355. doi:10.2307/1540861. ISSN0006-3185. JSTOR1540861.
↑ Drewett, Peter L. (2014-03-04), "Strombus gigas (Queen Conch)", Encyclopedia of Caribbean Archaeology, University Press of Florida, pp.329–330, doi:10.2307/j.ctvx1hst1.172
↑ Banaszak, Anastazia T.; García Ramos, Maribel; Goulet, Tamar L. (November 2013). "The symbiosis between the gastropod Strombus gigas and the dinoflagellate Symbiodinium: An ontogenic journey from mutualism to parasitism". Journal of Experimental Marine Biology and Ecology. 449: 358–365. Bibcode:2013JEMBE.449..358B. doi:10.1016/j.jembe.2013.10.027. ISSN0022-0981.
↑ BURGHARDT, I (2008-03-27). "Symbiosis between Symbiodinium (Dinophyceae) and various taxa of Nudibranchia (Mollusca: Gastropoda), with analyses of long-term retention". Organisms Diversity & Evolution. 8 (1): 66–76. Bibcode:2008ODivE...8...66B. doi:10.1016/j.ode.2007.01.001. ISSN1439-6092.
↑ Marco, Adolfo; Blaustein, Andrew R. (December 2000). "Symbiosis with Green Algae Affects Survival and Growth of Northwestern Salamander Embryos". Journal of Herpetology. 34 (4): 617. doi:10.2307/1565283. hdl:10261/48328. JSTOR1565283.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.