A DNA virus is a virus that has a genome made of deoxyribonucleic acid (DNA) that is replicated by a DNA polymerase. They can be divided between those that have two strands of DNA in their genome, called double-stranded DNA (dsDNA) viruses, and those that have one strand of DNA in their genome, called single-stranded DNA (ssDNA) viruses. dsDNA viruses primarily belong to two realms: Duplodnaviria and Varidnaviria, and ssDNA viruses are almost exclusively assigned to the realm Monodnaviria, which also includes some dsDNA viruses. Additionally, many DNA viruses are unassigned to higher taxa. Reverse transcribing viruses, which have a DNA genome that is replicated through an RNA intermediate by a reverse transcriptase, are classified into the kingdom Pararnavirae in the realm Riboviria.
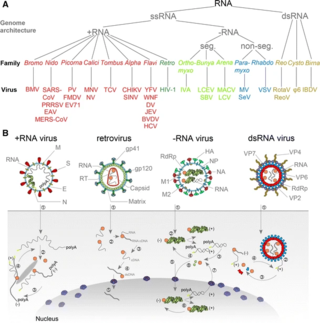
An RNA virus is a virus characterized by a ribonucleic acid (RNA) based genome. The genome can be single-stranded RNA (ssRNA) or double-stranded (dsRNA). Notable human diseases caused by RNA viruses include influenza, SARS, MERS, COVID-19, Dengue virus, hepatitis C, hepatitis E, West Nile fever, Ebola virus disease, rabies, polio, mumps, and measles.
Virus classification is the process of naming viruses and placing them into a taxonomic system similar to the classification systems used for cellular organisms.

Parvoviruses are a family of animal viruses that constitute the family Parvoviridae. They have linear, single-stranded DNA (ssDNA) genomes that typically contain two genes encoding for a replication initiator protein, called NS1, and the protein the viral capsid is made of. The coding portion of the genome is flanked by telomeres at each end that form into hairpin loops that are important during replication. Parvovirus virions are small compared to most viruses, at 23–28 nanometers in diameter, and contain the genome enclosed in an icosahedral capsid that has a rugged surface.

Geminiviridae is a family of plant viruses that encode their genetic information on a circular genome of single-stranded (ss) DNA. There are 520 species in this family, assigned to 14 genera. Diseases associated with this family include: bright yellow mosaic, yellow mosaic, yellow mottle, leaf curling, stunting, streaks, reduced yields. They have single-stranded circular DNA genomes encoding genes that diverge in both directions from a virion strand origin of replication. According to the Baltimore classification they are considered class II viruses. It is the largest known family of single stranded DNA viruses.
Baltimore classification is a system used to classify viruses based on their manner of messenger RNA (mRNA) synthesis. By organizing viruses based on their manner of mRNA production, it is possible to study viruses that behave similarly as a distinct group. Seven Baltimore groups are described that take into consideration whether the viral genome is made of deoxyribonucleic acid (DNA) or ribonucleic acid (RNA), whether the genome is single- or double-stranded, and whether the sense of a single-stranded RNA genome is positive or negative.

RNA-dependent RNA polymerase (RdRp) or RNA replicase is an enzyme that catalyzes the replication of RNA from an RNA template. Specifically, it catalyzes synthesis of the RNA strand complementary to a given RNA template. This is in contrast to typical DNA-dependent RNA polymerases, which all organisms use to catalyze the transcription of RNA from a DNA template.
Plectrovirus is a genus of viruses, in the family Plectroviridae. Bacteria in the phylum Mycoplasmatota serve as natural hosts, making these viruses bacteriophages. Acholeplasma virus L51 is the only species in the genus.

Bidensovirus is a genus of single stranded DNA viruses that infect invertebrates. The species in this genus were originally classified in the family Parvoviridae but were moved to a new genus because of significant differences in the genomes.
Spiraviridae is a family of incertae sedis viruses that replicate in hyperthermophilic archaea of the genus Aeropyrum, specifically Aeropyrum pernix. The family contains one genus, Alphaspiravirus, which contains one species, Aeropyrum coil-shaped virus. The virions of ACV are non-enveloped and in the shape of hollow cylinders that are formed by a coiling fiber that consists of two intertwining halves of the circular DNA strand inside a capsid. An appendage protrudes from each end of the cylindrical virion. The viral genome is ssDNA(+) and encodes for significantly more genes than other known ssDNA viruses. ACV is also unique in that it appears to lack its own enzymes to aid replication, instead likely using the host cell's replisomes. ACV has no known relation to any other archaea-infecting viruses, but it does share its coil-like morphology with some other archaeal viruses, suggesting that such viruses may be an ancient lineage that only infect archaea.

Positive-strand RNA viruses are a group of related viruses that have positive-sense, single-stranded genomes made of ribonucleic acid. The positive-sense genome can act as messenger RNA (mRNA) and can be directly translated into viral proteins by the host cell's ribosomes. Positive-strand RNA viruses encode an RNA-dependent RNA polymerase (RdRp) which is used during replication of the genome to synthesize a negative-sense antigenome that is then used as a template to create a new positive-sense viral genome.
Riboviria is a realm of viruses that includes all viruses that use a homologous RNA-dependent polymerase for replication. It includes RNA viruses that encode an RNA-dependent RNA polymerase, as well as reverse-transcribing viruses that encode an RNA-dependent DNA polymerase. RNA-dependent RNA polymerase (RdRp), also called RNA replicase, produces RNA from RNA. RNA-dependent DNA polymerase (RdDp), also called reverse transcriptase (RT), produces DNA from RNA. These enzymes are essential for replicating the viral genome and transcribing viral genes into messenger RNA (mRNA) for translation of viral proteins.
In virology, realm is the highest taxonomic rank established for viruses by the International Committee on Taxonomy of Viruses (ICTV), which oversees virus taxonomy. Six virus realms are recognized and united by specific highly conserved traits:

Smacoviridae is a family of single-stranded DNA viruses. The genomes of this family are small. The name Smacoviridae stands for 'small circular genome virus'. The genomes are circular single-stranded DNA and encode rolling-circle replication initiation proteins (Rep) and unique capsid proteins. As of 2021, 12 genera and 84 species are recognized in this family. The viruses in this taxon were isolated from faecal samples from insects and vertebrates by metagenomic methods. Little is known about their biology.

HUH endonucleases (HUH-tags) are sequence-specific single-stranded DNA (ssDNA) binding proteins originating from numerous species of bacteria and viruses. Viral HUH endonucleases are involved in initiating rolling circle replication while ones of bacterial origin initiate bacterial conjugation. In biotechnology, they can be used to create protein-DNA linkages, akin to other methods such as SNAP-tag. In doing so, they create a 5' covalent bond between the ssDNA and the protein. HUH endonucleases can be fused with other proteins or used as protein tags.
Cressdnaviricota is a phylum of viruses with small, circular single-stranded DNA genomes and encoding rolling circle replication-initiation proteins with the N-terminal HUH endonuclease and C-terminal superfamily 3 helicase domains. While the replication-associated proteins are homologous among viruses within the phylum, the capsid proteins are very diverse and have presumably been acquired from RNA viruses on multiple independent occasions. Nevertheless, all cressdnaviruses for which structural information is available appear to contain the jelly-roll fold.

Varidnaviria is a realm of viruses that includes all DNA viruses that encode major capsid proteins that contain a vertical jelly roll fold. The major capsid proteins (MCP) form into pseudohexameric subunits of the viral capsid, which stores the viral deoxyribonucleic acid (DNA), and are perpendicular, or vertical, to the surface of the capsid. Apart from this, viruses in the realm also share many other characteristics, such as minor capsid proteins (mCP) with the vertical jelly roll fold, an ATPase that packages viral DNA into the capsid, and a DNA polymerase that replicates the viral genome.

Bacilladnaviridae is a family of single-stranded DNA viruses that primarily infect diatoms.

Orthornavirae is a kingdom of viruses that have genomes made of ribonucleic acid (RNA), including genes which encode an RNA-dependent RNA polymerase (RdRp). The RdRp is used to transcribe the viral RNA genome into messenger RNA (mRNA) and to replicate the genome. Viruses in this kingdom share a number of characteristics which promote rapid evolution, including high rates of genetic mutation, recombination, and reassortment.
Rolling hairpin replication (RHR) is a unidirectional, strand displacement form of DNA replication used by parvoviruses, a group of viruses that constitute the family Parvoviridae. Parvoviruses have linear, single-stranded DNA (ssDNA) genomes in which the coding portion of the genome is flanked by telomeres at each end that form hairpin loops. During RHR, these hairpin loops repeatedly unfold and refold to change the direction of DNA replication so that replication progresses in a continuous manner back and forth across the genome. RHR is initiated and terminated by an endonuclease encoded by parvoviruses that is variously called NS1 or Rep, and RHR is similar to rolling circle replication, which is used by ssDNA viruses that have circular genomes.