Related Research Articles

A motor neuron is a neuron whose cell body is located in the motor cortex, brainstem or the spinal cord, and whose axon (fiber) projects to the spinal cord or outside of the spinal cord to directly or indirectly control effector organs, mainly muscles and glands. There are two types of motor neuron – upper motor neurons and lower motor neurons. Axons from upper motor neurons synapse onto interneurons in the spinal cord and occasionally directly onto lower motor neurons. The axons from the lower motor neurons are efferent nerve fibers that carry signals from the spinal cord to the effectors. Types of lower motor neurons are alpha motor neurons, beta motor neurons, and gamma motor neurons.

In neuroanatomy, the trigeminal nerve (lit. triplet nerve), also known as the fifth cranial nerve, cranial nerve V, or simply CN V, is a cranial nerve responsible for sensation in the face and motor functions such as biting and chewing; it is the most complex of the cranial nerves. Its name (trigeminal, from Latin tri- 'three', and -geminus 'twin') derives from each of the two nerves (one on each side of the pons) having three major branches: the ophthalmic nerve (V1), the maxillary nerve (V2), and the mandibular nerve (V3). The ophthalmic and maxillary nerves are purely sensory, whereas the mandibular nerve supplies motor as well as sensory (or "cutaneous") functions. Adding to the complexity of this nerve is that autonomic nerve fibers as well as special sensory fibers (taste) are contained within it.
In biology, a reflex, or reflex action, is an involuntary, unplanned sequence or action and nearly instantaneous response to a stimulus.

The somatic nervous system (SNS) is made up of nerves that link the brain and spinal cord to voluntary or skeletal muscles that are under conscious control as well as to skin sensory receptors. Specialized nerve fiber ends called sensory receptors are responsible for detecting information within and outside of the body.

Muscle spindles are stretch receptors within the body of a skeletal muscle that primarily detect changes in the length of the muscle. They convey length information to the central nervous system via afferent nerve fibers. This information can be processed by the brain as proprioception. The responses of muscle spindles to changes in length also play an important role in regulating the contraction of muscles, for example, by activating motor neurons via the stretch reflex to resist muscle stretch.

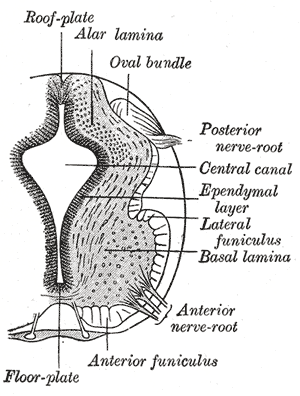
The grey column refers to a somewhat ridge-shaped mass of grey matter in the spinal cord. This presents as three columns: the anterior grey column, the posterior grey column, and the lateral grey column, all of which are visible in cross-section of the spinal cord.

A reflex arc is a neural pathway that controls a reflex. In vertebrates, most sensory neurons do not pass directly into the brain, but synapse in the spinal cord. This allows for faster reflex actions to occur by activating spinal motor neurons without the delay of routing signals through the brain. The brain will receive the input while the reflex is being carried out and the analysis of the signal takes place after the reflex action.
A mechanoreceptor, also called mechanoceptor, is a sensory receptor that responds to mechanical pressure or distortion. Mechanoreceptors are innervated by sensory neurons that convert mechanical pressure into electrical signals that, in animals, are sent to the central nervous system.
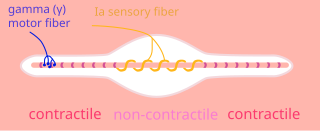
A nuclear chain fiber is a specialized sensory organ contained within a muscle. Nuclear chain fibers are intrafusal fibers that, along with nuclear bag fibers, make up the muscle spindle responsible for the detection of changes in muscle length.

Upper motor neurons (UMNs) is a term introduced by William Gowers in 1886. They are found in the cerebral cortex and brainstem and carry information down to activate interneurons and lower motor neurons, which in turn directly signal muscles to contract or relax. UMNs represent the major origin point for voluntary somatic movement.
A gamma motor neuron, also called gamma motoneuron, or fusimotor neuron, is a type of lower motor neuron that takes part in the process of muscle contraction, and represents about 30% of (Aγ) fibers going to the muscle. Like alpha motor neurons, their cell bodies are located in the anterior grey column of the spinal cord. They receive input from the reticular formation of the pons in the brainstem. Their axons are smaller than those of the alpha motor neurons, with a diameter of only 5 μm. Unlike the alpha motor neurons, gamma motor neurons do not directly adjust the lengthening or shortening of muscles. However, their role is important in keeping muscle spindles taut, thereby allowing the continued firing of alpha neurons, leading to muscle contraction. These neurons also play a role in adjusting the sensitivity of muscle spindles.

The vestibulospinal tract is a neural tract in the central nervous system. Specifically, it is a component of the extrapyramidal system and is classified as a component of the medial pathway. Like other descending motor pathways, the vestibulospinal fibers of the tract relay information from nuclei to motor neurons. The vestibular nuclei receive information through the vestibulocochlear nerve about changes in the orientation of the head. The nuclei relay motor commands through the vestibulospinal tract. The function of these motor commands is to alter muscle tone, extend, and change the position of the limbs and head with the goal of supporting posture and maintaining balance of the body and head.

The stretch reflex, or more accurately "muscle stretch reflex", is a muscle contraction in response to stretching a muscle. The function of the reflex is generally thought to be maintaining the muscle at a constant length but the response is often coordinated across multiple muscles and even joints. The older term deep tendon reflex is now criticized as misleading. Tendons have little to do with the response, and some muscles with stretch reflexes have no tendons. Rather, muscle spindles detect a stretch and convey the information to the central nervous system.
Alpha (α) motor neurons (also called alpha motoneurons), are large, multipolar lower motor neurons of the brainstem and spinal cord. They innervate extrafusal muscle fibers of skeletal muscle and are directly responsible for initiating their contraction. Alpha motor neurons are distinct from gamma motor neurons, which innervate intrafusal muscle fibers of muscle spindles.
Clasp-knife response refers to a Golgi tendon reflex with a rapid decrease in resistance when attempting to flex a joint, usually during a neurological examination. It is one of the characteristic responses of an upper motor neuron lesion. It gets its name from the resemblance between the motion of the limb and the sudden closing of a claspknife after sufficient pressure is applied.
Group A nerve fibers are one of the three classes of nerve fiber as generally classified by Erlanger and Gasser. The other two classes are the group B nerve fibers, and the group C nerve fibers. Group A are heavily myelinated, group B are moderately myelinated, and group C are unmyelinated.

A spinal interneuron, found in the spinal cord, relays signals between (afferent) sensory neurons, and (efferent) motor neurons. Different classes of spinal interneurons are involved in the process of sensory-motor integration. Most interneurons are found in the grey column, a region of grey matter in the spinal cord.

Cutaneous, superficial, or skin reflexes, are activated by skin receptors and play a valuable role in locomotion, providing quick responses to unexpected environmental challenges. They have been shown to be important in responses to obstacles or stumbling, in preparing for visually challenging terrain, and for assistance in making adjustments when instability is introduced. In addition to the role in normal locomotion, cutaneous reflexes are being studied for their potential in enhancing rehabilitation therapy (physiotherapy) for people with gait abnormalities.

The Golgi tendon organ (GTO) is a proprioceptor – a type of sensory receptor that senses changes in muscle tension. It lies at the interface between a muscle and its tendon known as the musculotendinous junction also known as the myotendinous junction. It provides the sensory component of the Golgi tendon reflex.
Proprioception refers to the sensory information relayed from muscles, tendons, and skin that allows for the perception of the body in space. This feedback allows for more fine control of movement. In the brain, proprioceptive integration occurs in the somatosensory cortex, and motor commands are generated in the motor cortex. In the spinal cord, sensory and motor signals are integrated and modulated by motor neuron pools called central pattern generators (CPGs). At the base level, sensory input is relayed by muscle spindles in the muscle and Golgi tendon organs (GTOs) in tendons, alongside cutaneous sensors in the skin.
References
- 1 2 3 Hall & Guyton (2006) , Golgi Tendon Reflex, pp. 679–680
- 1 2 3 4 Barrett et al (2010) , INVERSE STRETCH REFLEX, pp. 162–163
- 1 2 Saladin (2018) , The Tendon Reflex, pp. 498–499
- ↑ Michael-Titus, Adina T; Revest, Patricia; Shortland, Peter, eds. (2010). "Chapter 9 - Descending Pathways and Cerebellum". Systems of The Body: The Nervous System – Basic Science and Clinical Conditions (2nd ed.). Churchill Livingstone. Golgi tendon organs, p. 166. ISBN 9780702033735.
- 1 2 3 4 5 Pearson & Gordon (2013) , Convergence of Inputs on Ib Interneurons Increases the Flexibility of Reflex Responses, p. 799
- 1 2 3 Purves et al (2018a) , The Spinal Cord Circuitry Underlying the Regulation of Muscle Force, pp. 370–371
- ↑ Tortora, Gerard (2011). Principles of anatomy and physiology. Hoboken, N.J: Wiley. ISBN 9780470646083.
- ↑ Sharman MJ, Cresswell AG, Riek S (2006). "Proprioceptive neuromuscular facilitation stretching : mechanisms and clinical implications". Sports Med. 36 (11): 929–39. doi:10.2165/00007256-200636110-00002. PMID 17052131. S2CID 3123371.
- ↑ Pearson & Gordon (2013) , Figure 35–7 The reflex actions of Ib afferent fibers from Golgi tendon organs, p. 801
- ↑ Purves et al (2018b) , Box 17D Muscle Tone, p. 404
Other references
- Barrett, Kim E; Boitano, Scott; Barman, Susan M; Brooks, Heddwen L (2010). "Chapter 9 – Reflexes". Ganong's Review of Medical Physiology (23rd ed.). McGraw-Hill. ISBN 978-0-07-160567-0.
- Hall, JE; Guyton, AC (2006). "Chapter 54 - Motor Functions of the Spinal Cord; the Cord Reflexes". Textbook of medical physiology (11th ed.). St. Louis, Mo: Elsevier Saunders. pp. 673–684. ISBN 0-7216-0240-1.
- Pearson, Keir G; Gordon, James E (2013). "35 - Spinal Reflexes". In Kandel, Eric R; Schwartz, James H; Jessell, Thomas M; Siegelbaum, Steven A; Hudspeth, AJ (eds.). Principles of Neural Science (5th ed.). United States: McGraw-Hill. ISBN 978-0-07-139011-8.
- Purves, Dale; Augustine, George J; Fitzpatrick, David; Hall, William C; Lamantia, Anthony Samuel; Mooney, Richard D; Platt, Michael L; White, Leonard E, eds. (2018a). "Chapter 16 - Lower Motor Neuron Circuits and Motor Control". Neuroscience (6th ed.). Sinauer Associates. pp. 357–379. ISBN 9781605353807.
- Purves, Dale; Augustine, George J; Fitzpatrick, David; Hall, William C; Lamantia, Anthony Samuel; Mooney, Richard D; Platt, Michael L; White, Leonard E, eds. (2018b). "Chapter 17 - Upper Motor Neuron Control of the Brainstem and Spinal Cord". Neuroscience (6th ed.). Sinauer Associates. pp. 381–406. ISBN 9781605353807.
- Saladin, KS (2018). "Chapter 13 - The Spinal Cord, Spinal Nerves, and Somatic Reflexes". Anatomy and Physiology: The Unity of Form and Function (8th ed.). New York: McGraw-Hill. ISBN 978-1-259-27772-6.