Related Research Articles
Developmental biology is the study of the process by which animals and plants grow and develop. Developmental biology also encompasses the biology of regeneration, asexual reproduction, metamorphosis, and the growth and differentiation of stem cells in the adult organism.

Embryo drawing is the illustration of embryos in their developmental sequence. In plants and animals, an embryo develops from a zygote, the single cell that results when an egg and sperm fuse during fertilization. In animals, the zygote divides repeatedly to form a ball of cells, which then forms a set of tissue layers that migrate and fold to form an early embryo. Images of embryos provide a means of comparing embryos of different ages, and species. To this day, embryo drawings are made in undergraduate developmental biology lessons.
The theory of recapitulation, also called the biogenetic law or embryological parallelism—often expressed using Ernst Haeckel's phrase "ontogeny recapitulates phylogeny"—is an historical hypothesis that the development of the embryo of an animal, from fertilization to gestation or hatching (ontogeny), goes through stages resembling or representing successive adult stages in the evolution of the animal's remote ancestors (phylogeny). It was formulated in the 1820s by Étienne Serres based on the work of Johann Friedrich Meckel, after whom it is also known as Meckel–Serres law.

Ontogeny is the origination and development of an organism, usually from the time of fertilization of the egg to adult. The term can also be used to refer to the study of the entirety of an organism's lifespan.

Evolutionary developmental biology is a field of biological research that compares the developmental processes of different organisms to infer how developmental processes evolved.

In biology, homology is similarity due to shared ancestry between a pair of structures or genes in different taxa. A common example of homologous structures is the forelimbs of vertebrates, where the wings of bats and birds, the arms of primates, the front flippers of whales, and the forelegs of four-legged vertebrates like dogs and crocodiles are all derived from the same ancestral tetrapod structure. Evolutionary biology explains homologous structures adapted to different purposes as the result of descent with modification from a common ancestor. The term was first applied to biology in a non-evolutionary context by the anatomist Richard Owen in 1843. Homology was later explained by Charles Darwin's theory of evolution in 1859, but had been observed before this, from Aristotle onwards, and it was explicitly analysed by Pierre Belon in 1555.
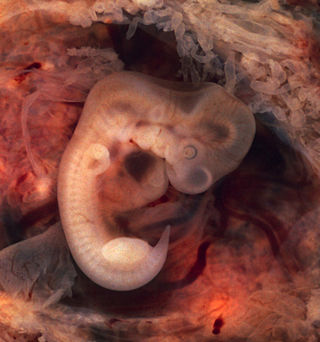
In biology, an atavism is a modification of a biological structure whereby an ancestral genetic trait reappears after having been lost through evolutionary change in previous generations. Atavisms can occur in several ways, one of which is when genes for previously existing phenotypic features are preserved in DNA, and these become expressed through a mutation that either knocks out the dominant genes for the new traits or makes the old traits dominate the new one. A number of traits can vary as a result of shortening of the fetal development of a trait (neoteny) or by prolongation of the same. In such a case, a shift in the time a trait is allowed to develop before it is fixed can bring forth an ancestral phenotype. Atavisms are often seen as evidence of evolution.

In evolutionary developmental biology, heterochrony is any genetically controlled difference in the timing, rate, or duration of a developmental process in an organism compared to its ancestors or other organisms. This leads to changes in the size, shape, characteristics and even presence of certain organs and features. It is contrasted with heterotopy, a change in spatial positioning of some process in the embryo, which can also create morphological innovation. Heterochrony can be divided into intraspecific heterochrony, variation within a species, and interspecific heterochrony, phylogenetic variation, i.e. variation of a descendant species with respect to an ancestral species.

Pleiotropy occurs when one gene influences two or more seemingly unrelated phenotypic traits. Such a gene that exhibits multiple phenotypic expression is called a pleiotropic gene. Mutation in a pleiotropic gene may have an effect on several traits simultaneously, due to the gene coding for a product used by a myriad of cells or different targets that have the same signaling function.
Hox genes, a subset of homeobox genes, are a group of related genes that specify regions of the body plan of an embryo along the head-tail axis of animals. Hox proteins encode and specify the characteristics of 'position', ensuring that the correct structures form in the correct places of the body. For example, Hox genes in insects specify which appendages form on a segment, and Hox genes in vertebrates specify the types and shape of vertebrae that will form. In segmented animals, Hox proteins thus confer segmental or positional identity, but do not form the actual segments themselves.

The theory of facilitated variation demonstrates how seemingly complex biological systems can arise through a limited number of regulatory genetic changes, through the differential re-use of pre-existing developmental components. The theory was presented in 2005 by Marc W. Kirschner and John C. Gerhart.

Phenotypic plasticity refers to some of the changes in an organism's behavior, morphology and physiology in response to a unique environment. Fundamental to the way in which organisms cope with environmental variation, phenotypic plasticity encompasses all types of environmentally induced changes that may or may not be permanent throughout an individual's lifespan.
In biology, saltation is a sudden and large mutational change from one generation to the next, potentially causing single-step speciation. This was historically offered as an alternative to Darwinism. Some forms of mutationism were effectively saltationist, implying large discontinuous jumps.
The pharyngula is a stage in the embryonic development of vertebrates. At this stage, the embryos of all vertebrates are similar, having developed features typical of vertebrates, such as the beginning of a spinal cord. Named by William Ballard, the pharyngula stage follows the blastula, gastrula and neurula stages.
Genetic assimilation is a process described by Conrad H. Waddington by which a phenotype originally produced in response to an environmental condition, such as exposure to a teratogen, later becomes genetically encoded via artificial selection or natural selection. Despite superficial appearances, this does not require the (Lamarckian) inheritance of acquired characters, although epigenetic inheritance could potentially influence the result. Waddington stated that genetic assimilation overcomes the barrier to selection imposed by what he called canalization of developmental pathways; he supposed that the organism's genetics evolved to ensure that development proceeded in a certain way regardless of normal environmental variations.

Ontogeny and Phylogeny is a 1977 book on evolution by Stephen Jay Gould, in which the author explores the relationship between embryonic development (ontogeny) and biological evolution (phylogeny). Unlike his many popular books of essays, it was a technical book, and over the following decades it was influential in stimulating research into heterochrony, which had been neglected since Ernst Haeckel's theory that ontogeny recapitulates phylogeny had been largely discredited. This helped to create the field of evolutionary developmental biology.

Acromelanism or colorpoint is a genetically determined, temperature-dependent pigmentation pattern, with full expression only occurring on legs, ears, tail, face and scrotum. It is most recognized as the coloration of Siamese and related breeds of cat, but can be found in dogs, rabbits, rats, mice, sheep, guinea pigs, minks, and gerbils. It is a specific type of point coloration.
In embryology a phylotypic stage or phylotypic period is a particular developmental stage or developmental period during mid-embryogenesis where embryos of related species within a phylum express the highest degree of morphological and molecular resemblance. Recent molecular studies in various plant and animal species were able to quantify the expression of genes covering crucial stages of embryo development and found that during the morphologically defined phylotypic period the evolutionary oldest genes, genes with similar temporal expression patterns, and genes under strongest purifying selection are most active throughout the phylotypic period.
In evolutionary biology, developmental bias refers to the production against or towards certain ontogenetic trajectories which ultimately influence the direction and outcome of evolutionary change by affecting the rates, magnitudes, directions and limits of trait evolution. Historically, the term was synonymous with developmental constraint, however, the latter has been more recently interpreted as referring solely to the negative role of development in evolution.
Ecological evolutionary developmental biology (eco-evo-devo) is a field of biology combining ecology, developmental biology and evolutionary biology to examine their relationship. The concept is closely tied to multiple biological mechanisms. The effects of eco-evo-devo can be a result of developmental plasticity, the result of symbiotic relationships or epigenetically inherited. The overlap between developmental plasticity and symbioses rooted in evolutionary concepts defines ecological evolutionary developmental biology. Host- microorganisms interactions during development characterize symbiotic relationships, whilst the spectrum of phenotypes rooted in canalization with response to environmental cues highlights plasticity. Developmental plasticity that is controlled by environmental temperature may put certain species at risk as a result of climate change.
References
- ↑ Zelditch, Miriam L.; Fink, William L. (2015). "Heterochrony and heterotopy: stability and innovation in the evolution of form". Paleobiology. 22 (2): 241–254. doi:10.1017/S0094837300016195. S2CID 89098289.
- 1 2 Held, Lewis I. (2014). How the Snake Lost its Legs. Curious Tales from the Frontier of Evo-Devo. Cambridge University Press. p. 152. ISBN 978-1-107-62139-8.
- ↑ Compagnucci, Claudia; Debiais-Thibaud, Melanie; Coolen, Marion; Fish, Jennifer; Griffin, John N.; Bertocchini, Federica; Minoux, Maryline; Rijli, Filippo M.; Borday-Birraux, Véronique; Casane, Didier; Mazan, Sylvie; Depew, Michael J. (2013). "Pattern and polarity in the development and evolution of the gnathostome jaw: Both conservation and heterotopy in the branchial arches of the shark, Scyliorhinus canicula". Developmental Biology. 377 (2): 428–448. doi: 10.1016/j.ydbio.2013.02.022 . PMID 23473983.
- ↑ Swanson, Christina I.; Schwimmer, David B.; Barolo, Scott (2011). "Rapid Evolutionary Rewiring of a Structurally Constrained Eye Enhancer". Current Biology. 21 (14): 1186–1196. Bibcode:2011CBio...21.1186S. doi:10.1016/j.cub.2011.05.056. PMC 3143281 . PMID 21737276.
- ↑ Hall, Brian K. (1999). "Time and Place in Evolution: Heterochrony and Heterotopy". Evolutionary Developmental Biology. pp. 375–391. doi:10.1007/978-94-011-3961-8_24. ISBN 978-0-412-78590-0.