Non-homologous end joining (NHEJ) is a pathway that repairs double-strand breaks in DNA. NHEJ is referred to as "non-homologous" because the break ends are directly ligated without the need for a homologous template, in contrast to homology directed repair, which requires a homologous sequence to guide repair. The term "non-homologous end joining" was coined in 1996 by Moore and Haber.

The T-cell receptor (TCR) is a protein complex found on the surface of T cells, or T lymphocytes, that is responsible for recognizing fragments of antigen as peptides bound to major histocompatibility complex (MHC) molecules. The binding between TCR and antigen peptides is of relatively low affinity and is degenerate: that is, many TCRs recognize the same antigen peptide and many antigen peptides are recognized by the same TCR.
In the human immune system, central tolerance is the process of eliminating any developing T or B lymphocytes that are reactive to self. Through elimination of autoreactive lymphocytes, tolerance ensures that the immune system does not attack self peptides. Lymphocyte maturation occurs in primary lymphoid organs such as the bone marrow and the thymus. In mammals, B cells mature in the bone marrow and T cells mature in the thymus.

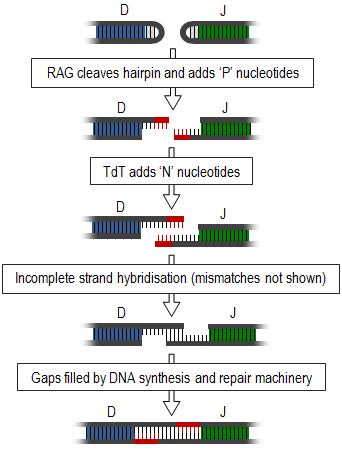
Terminal deoxynucleotidyl transferase (TdT), also known as DNA nucleotidylexotransferase (DNTT) or terminal transferase, is a specialized DNA polymerase expressed in immature, pre-B, pre-T lymphoid cells, and acute lymphoblastic leukemia/lymphoma cells. TdT adds N-nucleotides to the V, D, and J exons of the TCR and BCR genes during antibody gene recombination, enabling the phenomenon of junctional diversity. In humans, terminal transferase is encoded by the DNTT gene. As a member of the X family of DNA polymerase enzymes, it works in conjunction with polymerase λ and polymerase μ, both of which belong to the same X family of polymerase enzymes. The diversity introduced by TdT has played an important role in the evolution of the vertebrate immune system, significantly increasing the variety of antigen receptors that a cell is equipped with to fight pathogens. Studies using TdT knockout mice have found drastic reductions (10-fold) in T-cell receptor (TCR) diversity compared with that of normal, or wild-type, systems. The greater diversity of TCRs that an organism is equipped with leads to greater resistance to infection. Although TdT was one of the first DNA polymerases identified in mammals in 1960, it remains one of the least understood of all DNA polymerases. In 2016–18, TdT was discovered to demonstrate in trans template dependant behaviour in addition to its more broadly known template independent behaviour
V(D)J recombination is the mechanism of somatic recombination that occurs only in developing lymphocytes during the early stages of T and B cell maturation. It results in the highly diverse repertoire of antibodies/immunoglobulins and T cell receptors (TCRs) found in B cells and T cells, respectively. The process is a defining feature of the adaptive immune system.

A palindromic sequence is a nucleic acid sequence in a double-stranded DNA or RNA molecule whereby reading in a certain direction on one strand is identical to the sequence in the same direction on the complementary strand. This definition of palindrome thus depends on complementary strands being palindromic of each other.
Allelic exclusion is a process by which only one allele of a gene is expressed while the other allele is silenced. This phenomenon is most notable for playing a role in the development of B lymphocytes, where allelic exclusion allows for each mature B lymphocyte to express only one type of immunoglobulin. This subsequently results in each B lymphocyte being able to recognize only one antigen. This is significant as the co-expression of both alleles in B lymphocytes is associated with autoimmunity and the production of autoantibodies.
The recombination-activating genes (RAGs) encode parts of a protein complex that plays important roles in the rearrangement and recombination of the genes encoding immunoglobulin and T cell receptor molecules. There are two recombination-activating genes RAG1 and RAG2, whose cellular expression is restricted to lymphocytes during their developmental stages. The enzymes encoded by these genes, RAG-1 and RAG-2, are essential to the generation of mature B cells and T cells, two types of lymphocyte that are crucial components of the adaptive immune system.

DNA repair protein XRCC4 also known as X-ray repair cross-complementing protein 4 or XRCC4 is a protein that in humans is encoded by the XRCC4 gene. In addition to humans, the XRCC4 protein is also expressed in many other metazoans, fungi and in plants. The X-ray repair cross-complementing protein 4 is one of several core proteins involved in the non-homologous end joining (NHEJ) pathway to repair DNA double strand breaks (DSBs).

Immunoglobulin class switching, also known as isotype switching, isotypic commutation or class-switch recombination (CSR), is a biological mechanism that changes a B cell's production of immunoglobulin from one type to another, such as from the isotype IgM to the isotype IgG. During this process, the constant-region portion of the antibody heavy chain is changed, but the variable region of the heavy chain stays the same. Since the variable region does not change, class switching does not affect antigen specificity. Instead, the antibody retains affinity for the same antigens, but can interact with different effector molecules.

In immunology, an idiotype is a shared characteristic between a group of immunoglobulin or T-cell receptor (TCR) molecules based upon the antigen binding specificity and therefore structure of their variable region. The variable region of antigen receptors of T cells (TCRs) and B cells (immunoglobulins) contain complementarity-determining regions (CDRs) with unique amino acid sequences. They define the surface and properties of the variable region, determining the antigen specificity and therefore the idiotope of the molecule. Immunoglobulins or TCRs with a shared idiotope are the same idiotype. Antibody idiotype is determined by:

Artemis is a protein that in humans is encoded by the DCLRE1C gene.
DNA polymerase lambda, also known as Pol λ, is an enzyme found in all eukaryotes. In humans, it is encoded by the POLL gene.
T cell receptor gamma locus is a protein that in humans is encoded by the TRG gene, also known as TCRG or TRG@. It contributes the gamma (γ) chain to the larger TCR protein.

DNA polymerase mu is a polymerase enzyme found in eukaryotes. In humans, this protein is encoded by the POLM gene.
Recombination signal sequences are conserved sequences of noncoding DNA that are recognized by the RAG1/RAG2 enzyme complex during V(D)J recombination in immature B cells and T cells. Recombination signal sequences guide the enzyme complex to the V, D, and J gene segments that will undergo recombination during the formation of the heavy and light-chain variable regions in T-cell receptors and immunoglobulin molecules.
Somatic hypermutation is a cellular mechanism by which the immune system adapts to the new foreign elements that confront it, as seen during class switching. A major component of the process of affinity maturation, SHM diversifies B cell receptors used to recognize foreign elements (antigens) and allows the immune system to adapt its response to new threats during the lifetime of an organism. Somatic hypermutation involves a programmed process of mutation affecting the variable regions of immunoglobulin genes. Unlike germline mutation, SHM affects only an organism's individual immune cells, and the mutations are not transmitted to the organism's offspring. Mistargeted somatic hypermutation is a likely mechanism in the development of B-cell lymphomas and many other cancers.
The Artemis complex is a protein complex that functions in V(D)J recombination, the somatic recombination process which generates diversity in T cell receptors and immunoglobulins. Mutations in the Artemis complex results in hypersensitivity to DNA double-strand break-inducing agents, such as radiation; and so people with mutations in the Artemis complex may develop radiosensitive severe combined immune deficiency (RS-SCID).
Antibody structure is made up of two heavy-chains and two light-chains. These chains are held together by disulfide bonds. The arrangement or processes that put together different parts of this antibody molecule play important role in antibody diversity and production of different subclasses or classes of antibodies. The organization and processes take place during the development and differentiation of B cells. That is, the controlled gene expression during transcription and translation coupled with the rearrangements of immunoglobulin gene segments result in the generation of antibody repertoire during development and maturation of B cells.
T-cell receptor revision is a process in the peripheral immune system which is used by mature T cells to alter their original antigenic specificity based on rearranged T cell receptors (TCR). This process can lead either to continuous appearance of potentially self-reactive T cells in the body, not controlled by the central tolerance mechanism in the thymus or better eliminate such self-reactive T cells on the other hand and thus contributing to peripheral tolerance - the extent of each has not been completely understood yet. This process occurs during follicular helper T cell formation in lymph node germinal centers.