
A lichen is a hybrid colony of algae or cyanobacteria living symbiotically among filaments of multiple fungi species, along with a yeast embedded in the cortex or "skin", in a mutualistic relationship.

The Rhynie chert is a Lower Devonian sedimentary deposit exhibiting extraordinary fossil detail or completeness. It is exposed near the village of Rhynie, Aberdeenshire, Scotland; a second unit, the Windyfield chert, is located some 700 m away.

Baragwanathia is a genus of extinct lycopsid plants of Late Silurian to Early Devonian age, fossils of which have been found in Australia, Canada, China and Czechia. The name derives from William Baragwanath who discovered the first specimens of the type species, Baragwanathia longifolia, at Thomson River.

Protosalvinia is a prehistoric plant found commonly in shale from shoreline habitats of the Upper Devonian period. The name Protosalvinia is a misnomer. The name literally means early Salvinia, and was given in the erroneous belief that the fossils were an earlier form of the living aquatic fern Salvinia. It is no longer believed that the fossils come from a fern, but deciding exactly what the fossils represent is still a matter of debate.

Prototaxites is an extinct genus of terrestrial fungi dating from the Late Silurian until the Late Devonian periods. Prototaxites formed large trunk-like structures up to 1 metre (3 ft) wide, reaching 8 metres (26 ft) in length, made up of interwoven tubes around 50 micrometres (0.0020 in) in diameter, making it by far the largest land-dwelling organism of its time.

Cosmochlaina is a form genus of nematophyte – an early plant known only from fossil cuticles, often found in association with tubular structures. The form genus was put forwards by Dianne Edwards, and is diagnosed by inwards-pointing flanges and randomly oriented pseudo-cellular units. Projections on the outer surface are always present, and sometimes also appear on the inner surface; however, the surface of the cuticle itself is always smooth. The holes in the cuticle are often covered by round flaps, loosely attached along a side.
The Nematophyta or nematophytes are a paraphyletic group of land organisms, probably including some plants as well as algae known only from the fossil record, from the Silurian period until the early Devonian Rhynie chert. The type genus Nematothallus, which typifies the group, was first described by Lang in 1937, who envisioned it being a thallose plant with tubular features and sporophytes, covered by a cuticle which preserved impressions of the underlying cells. He had found abundant disaggregated remains of all three features, none of which were connected to another, leaving his reconstruction of the phytodebris as parts of a single organism highly conjectural. No reproductive or vegetative structures common to the land plants are known, and certain members of the nematophyte plexus seem to belong to the fungi.
Ornatifilum is an artificial form genus, which is used to categorise any small, branched filaments with external ornamentation.

Aglaophyton major was the sporophyte generation of a diplohaplontic, pre-vascular, axial, free-sporing land plant of the Lower Devonian. It had anatomical features intermediate between those of the bryophytes and vascular plants or tracheophytes.
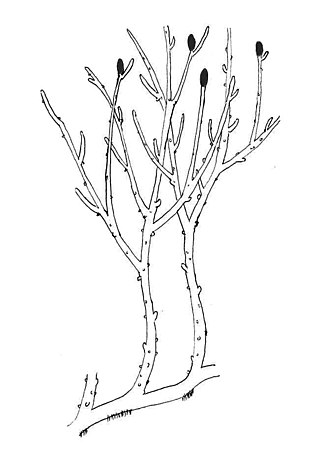
Rhynia is a single-species genus of Devonian vascular plants. Rhynia gwynne-vaughanii was the sporophyte generation of a vascular, axial, free-sporing diplohaplontic embryophytic land plant of the Early Devonian that had anatomical features more advanced than those of the bryophytes. Rhynia gwynne-vaughanii was a member of a sister group to all other eutracheophytes, including modern vascular plants.
This article attempts to place key plant innovations in a geological context. It concerns itself only with novel adaptations and events that had a major ecological significance, not those that are of solely anthropological interest. The timeline displays a graphical representation of the adaptations; the text attempts to explain the nature and robustness of the evidence.
Professor Dianne Edwards CBE, FRS, FRSE, FLS, FLSW is a palaeobotanist, who studies the colonisation of land by plants, and early land plant interactions.
Nematoplexus rhyniensis is a fossil known from the Rhynie chert assigned to the nematophytes. It comprises a loose mass of coily aseptate tubes. Tubes which may have originated from a Nematoplexus-like plant are known from earlier Silurian sediments.
Steganotheca is a genus of bushy, probably vascular plants with branched axes, known from upper Silurian strata. It has terminal sporangia and reached 5 cm in height.

Banded tubes are a type of phytoclast consisting of micrometre-scale tubes with a banded internal ornamentation, and known from terrestrial/freshwater settings from the Early Silurian onwards.
Nematasketum is a nematophyte with internally thickened tubes. It is thought to be terrestrial or freshwater, and seems to be aligned with the fungi.

Uskiella is a genus of small fossil plants of Early Devonian age. The diagnostic characters are naked axes branching isotomously, terminating in ellipsoidal, vertically elongate flat sporangia which split longitudinally into two valves. Spores of U. reticulata have a reticulate appearance. Coalified specimens have been reported from Wales, with a possible occurrence in Australia.
Grisellatheca was a genus of land plant with branching axes. It is known from charcoalified Early Devonian deposits, its type locality being the Brown Clee Hill lagerstatten. Its Terahedraletes spores form permanent tetrads.
Psilophytites is a form genus of extinct plants; it was created by Høeg for spiny stems (axes) which cannot be assigned to a more precise genus or species, usually because spore-forming organs or sporangia are not present.
Tortotubus is an early terrestrial fungus. Its growth trajectory can be ascertained from its fossils, which occur across the globe from the Ordovician to the Devonian. These fossils document foraging activities of slender, cell-wide exploratory hyphae; when these hit a source of food, they produced secondary branches that grew back down the original filament, covered themselves with an envelope, and served as pipes to shuttle nutrients to other parts of the organism. Today, mycelium with this growth pattern is observed in the mushroom-forming fungi.




