Paratrimastix pyriformis Last updated October 16, 2025 History of knowledge This species was first described by G. A. Klebs in 1892 as Tetramitus pyriformis . [ 2] [ 3] Under this name, it has been frequently discussed in the context of sewage , sewage treatment , and water quality during the 20th century. [ 4] [ 5] [ 6] [ 7] [ 8] [ 9] [ 10] [ 11] [ 12] [ 13] [ 14] It was also observed on Elephant Island , South Shetland Islands . [ 15] More than 100 years after its description, in 1999, it was transferred to the genus Trimastix based on its morphology . [ 16] The first ultrastructural study using transmission electron microscopy was published the same year, which reported a discovery of hydrogenosome -like organelles in the species. [ 17]
A molecular phylogenetic study based on small-subunit ribosomal RNA placed the genus Trimastix (then including P. pyriformis ) as sister to the oxymonad Pyrsonympha in 2001 [ 18] and a close relationship to oxymonads was further supported in another study in 2005. [ 19] The clade uniting Trimastix and oxymonads was named Preaxostyla in 2003. [ 20] A more detailed molecular phylogenetic analysis in 2015 placed this species in a new genus Paratrimastix , even more closely related to oxymonads than Trimastix . [ 21] Preaxostyla (consisting of Trimastix , Paratrimastix , and oxymonads ) is now considered one of the five major lineages of Metamonada . [ 22] [ 23]
The interest in P. pyriformis , and especially its reduced mitochondria, was largely driven by the possibility that oxymonads might be completely amitochondrial. [ 24] This was supported by a genomic analysis of Monocercomonoides exilis published in 2016, which demonstrated that this oxymonad is the first known eukaryote that has completely lost its mitochondria . [ 25] [ 26] [ 27]
A series of transcriptomic studies between the years 2006 and 2016 reported details of P. pyriformis glycolytic pathway [ 28] and arginine deiminase pathway, [ 29] as well as supported the mitochondrial ancestry of its hydrogenosome -like organelles [ 30] and uncovered their role in amino acid metabolism. [ 31] Preliminary results of a genomic project led to the characterization of the unusual preaxostylan type iron-sulfur cluster assembly machinery in P. pyriformis in 2018, [ 32] the role of its reduced mitochondria in the methionine cycle (2022), [ 33] and the experimental characterization of one of its mitochondrial carriers (2023). [ 34] The complete genomic assembly of P. pyriformis was published in 2023 in a large-scale comparative genomic study focused on the reductive evolution of mitochondria in Preaxostyla , which also identified two additional oxymonad species with no traces of mitochondria . [ 35]
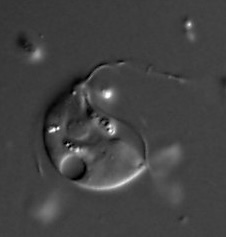
Morphology and ultrastructure Paratrimastix pyriformis has four flagella , one directed anteriorly, one posteriorly, and others laterally. The posterior flagellum has two vanes with thickened vane margins. Both vanes have a paracrystalline substructure. Ventral side of the cell is shaped in the form of a broad groove, a typical excavate feature, which is used as a feeding structure. The cells measure 9-17 μm in length and 5-13 μm in width. The single nucleus with a conspicuous central nucleolus is located in the anterior third of the cell. [ 17]
Dense network of rough endoplasmic reticulum extends from the nucleus towards the posterior end of the cell. A single stacked Golgi apparatus is located posterior and to the left of the basal bodies . The kinetid consists of four basal bodies , four microtubular roots, and various microtubules and fibers associated with the basal bodies and roots. The arrangement of the basal bodies is asymmetrical. Rod-shaped mitochondrion -related organelles resembling hydrogenosomes are 0.5-1.0 μm in length and bounded by a double membrane . The mitochondrion -related organelles are dispersed throughout the cell. [ 17]
Paratrimastix pyriformis may be distinguished from the marine Trimastix marina and the freshwater Paratrimastix eleionoma [ 1] by the non-thickened and discretely subapically inserting anterior flagellum , [ 16] from Trimastix inaequalis by the equal length of its lateral flagella , and from Trimastix convexa (most similar species) by its smaller size and ultrastructural details of the cytoskeleton . [ 17]
Behaviour Paratrimastix pyriformis swims with the anterior and lateral flagella beating and rotates occasionally. It can attach to the substrate by the tip of the posterior flagellum. Its cell contains food vacuoles with bacteria . A small contractile vacuole is located posteriorly. [ 16] Bacteria are captured at the posterior end of the ventral groove. Flagella are retained throughout cell division . Some, but not all, strains of P. pyriformis produce cysts: rounded cells with thin walls and basal bodies and flagella preserved. [ 17]
Glycolysis in P. pyriformis includes at least four alternative enzymes that have likely been gained by lateral gene transfer from Bacteria . [ 28] P. pyriformis further produces additional ATP using the extended glycolysis pathway where pyruvate generated in glycolysis is metabolised into acetate , CO2 , and H2 . Alternatively, pyruvate can be produced by decarboxylation of malate through the activity of the malic enzyme (ME). Pyruvate is decarboxylated to acetyl coenzyme A by pyruvate:ferredoxin oxidoreductase (PFO). The last part of the pathway, which yields ATP , acetate , and coenzyme A is catalyzed by a single enzyme: acetyl-CoA synthetase (ACS) like in the diplomonad Giardia intestinalis . Activities of both ME and PFO produce excess electrons which are then consumed in reduction of protons to molecular hydrogen through the activity of [FeFe] hydrogenases . [ 35]
Based on an in-silico reconstructed amino acid metabolism, P. pyriformis is able to synthesize at least five protein-forming amino acids including selenocysteine . [ 35] Unlike other Preaxostyla , P. pyriformis doesn't have a complete arginine deiminase pathway, [ 29] and therefore is likely unable to produce ATP via arginine catabolism . However, other amino acids (cysteine , serine , tryptophan , and methionine ) can hypothetically be utilized to produce ATP by their conversion of pyruvate and α-keto-butyrate , which can enter the extended glycolytic pathway. [ 35] [ 36]
References 1 2 Zhang, Qianqian; Táborský, Petr; Silberman, Jeffrey D.; Pánek, Tomáš; Čepička, Ivan; Simpson, Alastair G.B. (September 2015). "Marine Isolates of Trimastix marina Form a Plesiomorphic Deep-branching Lineage within Preaxostyla, Separate from Other Known Trimastigids (Paratrimastix n. gen.)" . Protist . 166 (4): 468– 491. doi :10.1016/j.protis.2015.07.003 . PMID 26312987 . ↑ "PID - Trimastix Taxonomy" . megasun.bch.umontreal.ca . Retrieved 2024-04-15 . ↑ "WoRMS - World Register of Marine Species" . www.marinespecies.org . Retrieved 2024-04-15 . ↑ Lackey, James B. (1932). "Oxygen Deficiency and Sewage Protozoa: With Descriptions of Some New Species" . The Biological Bulletin . 63 (2): 287– 295. doi :10.2307/1537245 . ISSN 0006-3185 . JSTOR 1537245 . ↑ Liebmann, Hans (1938). "Weitere Beiträge zur Kenntnis der Protozoenfauna des Faulschlammes der Bleilochsperre" . Archiv für Protistenkunde . 90 (1938): 272– 291. ↑ Lackey, James B. (1942). "The Effects of Distillery Wastes and Waters on the Microscopic Flora and Fauna of a Small Creek". Public Health Reports . 57 (8): 253– 260. doi :10.2307/4584018 . ISSN 0094-6214 . JSTOR 4584018 . ↑ Lackey, James B.; Dixon, R. M. (1943). "Some Biological Aspects of the Hays Process of Sewage Treatment". Sewage Works Journal . 15 (6): 1139– 1152. ISSN 0096-9362 . JSTOR 25029701 . ↑ Water Quality Measurement and Instrumentation: Proceedings ↑ Ingram, William T. (1962). Microbiological Waste Treatment Processes in a Closed Ecology ↑ Calaway, Wilson Thayer (1962). Waste treatment protozoa: Flagellata . Florida Univ., Gainesville. Engineering and Industrial Experiment Station. ISSN 0428-6863 . ↑ József, Hamar (1971). "Effect of the waste-water of sogar-works on natural history of the river Zagyva" . Tiscia (in Hungarian) (6): 109– 128. ISSN 0563-587X . ↑ Šimková, Mária; Dudová, Danka (1988). "Mikrobiologické hodnotenie aktivovaných kalov z čistiarní odpadových vôd vybraných potravinárskych závodov" . Bulletin potravinárskeho výskumu . 27 : 143– 148. ↑ Foissner, Wilhelm (1992). "Evaluating water quality using protozoa and saprobity indexes" . Protocols in Protozoology : B-11.1 – B-11.20. ↑ Berger, Helmut; Foissner, Wilhelm; Kohmann, Fritz (1997). Bestimmung und Ökologie der Mikrosaprobien nach DIN 38410: 26 Bildbestimmungsschlüssel, 10 Tabellen . Stuttgart Jena Lübeck Ulm: G. Fischer. ISBN 978-3-437-25026-2 ↑ Smith, H. G. (1972). "The terrestrial Protozoa of Elephant Island, South Shetland Islands" . British Antarctic Survey Bulletin . 31 : 55– 62. 1 2 3 Bernard, Catherine; Simpson, Alastair G. B.; Patterson, David J. (May 2000). "Some free-living flagellates (protista) from anoxic habitats" . Ophelia . 52 (2): 113– 142. doi :10.1080/00785236.1999.10409422 . ISSN 0078-5326 . 1 2 3 4 5 O'Kelly, Charles J.; Farmer, Mark A.; Nerad, Thomas A. (August 1999). "Ultrastructure of Trimastix pyriformis (Klebs) Bernard et al.: Similarities of Trimastix Species with Retortamonad and Jakobid Flagellates" . Protist . 150 (2): 149– 162. doi :10.1016/S1434-4610(99)70018-0 . PMID 10505415 . ↑ Dacks, Joel B.; Silberman, Jeffrey D.; Simpson, Alastair G. B.; Moriya, Shigeharu; Kudo, Toshiaki; Ohkuma, Moriya; Redfield, Rosemary J. (2001-06-01). "Oxymonads Are Closely Related to the Excavate Taxon Trimastix" . Molecular Biology and Evolution . 18 (6): 1034– 1044. doi :10.1093/oxfordjournals.molbev.a003875 . ISSN 1537-1719 . PMID 11371592 . ↑ Hampl, Vladimír; Horner, David S.; Dyal, Patricia; Kulda, Jaroslav; Flegr, Jaroslav; Foster, Peter G.; Embley, T. Martin (2005-12-01). "Inference of the Phylogenetic Position of Oxymonads Based on Nine Genes: Support for Metamonada and Excavata" . Molecular Biology and Evolution . 22 (12): 2508– 2518. doi :10.1093/molbev/msi245 . ISSN 1537-1719 . PMID 16120804 . ↑ Simpson, A. G. B. (2003-11-01). "Cytoskeletal organization, phylogenetic affinities and systematics in the contentious taxon Excavata (Eukaryota)" . International Journal of Systematic and Evolutionary Microbiology . 53 (6): 1759– 1777. doi :10.1099/ijs.0.02578-0 . ISSN 1466-5026 . PMID 14657103 . ↑ Zhang, Qianqian; Táborský, Petr; Silberman, Jeffrey D.; Pánek, Tomáš; Čepička, Ivan; Simpson, Alastair G.B. (September 2015). "Marine Isolates of Trimastix marina Form a Plesiomorphic Deep-branching Lineage within Preaxostyla, Separate from Other Known Trimastigids (Paratrimastix n. gen.)" . Protist . 166 (4): 468– 491. doi :10.1016/j.protis.2015.07.003 . PMID 26312987 . ↑ Stairs, Courtney W.; Táborský, Petr; Salomaki, Eric D.; Kolisko, Martin; Pánek, Tomáš; Eme, Laura; Hradilová, Miluše; Vlček, Čestmír; Jerlström-Hultqvist, Jon; Roger, Andrew J.; Čepička, Ivan (December 2021). "Anaeramoebae are a divergent lineage of eukaryotes that shed light on the transition from anaerobic mitochondria to hydrogenosomes" . Current Biology . 31 (24): 5605–5612.e5. Bibcode :2021CBio...31E5605S . doi :10.1016/j.cub.2021.10.010 . PMID 34710348 . ↑ Yazaki, Euki; Kume, Keitaro; Shiratori, Takashi; Eglit, Yana; Tanifuji, Goro; Harada, Ryo; Simpson, Alastair G. B.; Ishida, Ken-ichiro; Hashimoto, Tetsuo; Inagaki, Yuji (2020-09-09). "Barthelonids represent a deep-branching metamonad clade with mitochondrion-related organelles predicted to generate no ATP" . Proceedings of the Royal Society B: Biological Sciences . 287 (1934) 20201538. doi :10.1098/rspb.2020.1538 . ISSN 0962-8452 . PMC 7542792 . PMID 32873198 . ↑ Novak, Lukas (17 September 2013). "Mitochondrion of Trimastix pyriformis (diploma thesis)" . Charles University Digital Repository . ↑ Karnkowska, Anna; Vacek, Vojtěch; Zubáčová, Zuzana; Treitli, Sebastian C.; Petrželková, Romana; Eme, Laura; Novák, Lukáš; Žárský, Vojtěch; Barlow, Lael D.; Herman, Emily K.; Soukal, Petr; Hroudová, Miluše; Doležal, Pavel; Stairs, Courtney W.; Roger, Andrew J. (2016). "A Eukaryote without a Mitochondrial Organelle" . Current Biology . 26 (10): 1274– 1284. Bibcode :2016CBio...26.1274K . doi : 10.1016/j.cub.2016.03.053 . PMID 27185558 . ↑ Treitli, Sebastian Cristian; Peña-Diaz, Priscila; Hałakuc, Paweł; Karnkowska, Anna; Hampl, Vladimír (2021). "High quality genome assembly of the amitochondriate eukaryote Monocercomonoides exilis" . Microbial Genomics . 7 (12): 000745. doi : 10.1099/mgen.0.000745 . ISSN 2057-5858 . PMC 8767320 . PMID 34951395 . ↑ Karnkowska, Anna; Treitli, Sebastian C; Brzoň, Ondřej; Novák, Lukáš; Vacek, Vojtěch; Soukal, Petr; Barlow, Lael D; Herman, Emily K; Pipaliya, Shweta V; Pánek, Tomáš; Žihala, David; Petrželková, Romana; Butenko, Anzhelika; Eme, Laura; Stairs, Courtney W (2019-10-01). Battistuzzi, Fabia Ursula (ed.). "The Oxymonad Genome Displays Canonical Eukaryotic Complexity in the Absence of a Mitochondrion" . Molecular Biology and Evolution . 36 (10): 2292– 2312. doi :10.1093/molbev/msz147 . ISSN 0737-4038 . PMC 6759080 . PMID 31387118 . 1 2 Stechmann, Alexandra; Baumgartner, Manuela; Silberman, Jeffrey D; Roger, Andrew J (2006). "The glycolytic pathway of Trimastix pyriformis is an evolutionary mosaic" . BMC Evolutionary Biology . 6 (1): 101. doi : 10.1186/1471-2148-6-101 . PMC 1665464 . PMID 17123440 . 1 2 Novák, Lukáš; Zubáčová, Zuzana; Karnkowska, Anna; Kolisko, Martin; Hroudová, Miluše; Stairs, Courtney W.; Simpson, Alastair G. B.; Keeling, Patrick J.; Roger, Andrew J.; Čepička, Ivan; Hampl, Vladimír (December 2016). "Arginine deiminase pathway enzymes: evolutionary history in metamonads and other eukaryotes" . BMC Evolutionary Biology . 16 (1): 197. Bibcode :2016BMCEE..16..197N . doi : 10.1186/s12862-016-0771-4 . ISSN 1471-2148 . PMC 5052871 . PMID 27716026 . ↑ Hampl, Vladimir; Silberman, Jeffrey D.; Stechmann, Alexandra; Diaz-Triviño, Sara; Johnson, Patricia J.; Roger, Andrew J. (2008-01-02). Redfield, Rosemary (ed.). "Genetic Evidence for a Mitochondriate Ancestry in the 'Amitochondriate' Flagellate Trimastix pyriformis" . PLOS ONE . 3 (1) e1383. Bibcode :2008PLoSO...3.1383H . doi : 10.1371/journal.pone.0001383 . ISSN 1932-6203 . PMC 2148110 . PMID 18167542 . ↑ Zubáčová, Zuzana; Novák, Lukáš; Bublíková, Jitka; Vacek, Vojtěch; Fousek, Jan; Rídl, Jakub; Tachezy, Jan; Doležal, Pavel; Vlček, Čestmír; Hampl, Vladimír (2013-03-13). Saks, Valdur (ed.). "The Mitochondrion-Like Organelle of Trimastix pyriformis Contains the Complete Glycine Cleavage System" . PLOS ONE . 8 (3) e55417. Bibcode :2013PLoSO...855417Z . doi : 10.1371/journal.pone.0055417 . ISSN 1932-6203 . PMC 3596361 . PMID 23516392 . ↑ Vacek, Vojtěch; Novák, Lukáš V F; Treitli, Sebastian C; Táborský, Petr; Čepička, Ivan; Kolísko, Martin; Keeling, Patrick J; Hampl, Vladimír (2018-09-01). Ruiz-Trillo, Iñaki (ed.). "Fe–S Cluster Assembly in Oxymonads and Related Protists" . Molecular Biology and Evolution . 35 (11): 2712– 2718. doi :10.1093/molbev/msy168 . ISSN 0737-4038 . PMC 6231488 . PMID 30184127 . ↑ Zítek, Justyna; Füssy, Zoltán; Treitli, Sebastian C.; Peña-Diaz, Priscila; Vaitová, Zuzana; Zavadska, Daryna; Harant, Karel; Hampl, Vladimír (2022). "Reduced mitochondria provide an essential function for the cytosolic methionine cycle" . Current Biology . 32 (23): 5057–5068.e5. Bibcode :2022CBio...32E5057Z . doi :10.1016/j.cub.2022.10.028 . PMC 9746703 . PMID 36347252 . ↑ Zítek, Justyna; King, Martin S.; Peña-Diaz, Priscila; Pyrihová, Eva; King, Alannah C.; Kunji, Edmund R.S.; Hampl, Vladimír (2023). "The free-living flagellate Paratrimastix pyriformis uses a distinct mitochondrial carrier to balance adenine nucleotide pools" . Archives of Biochemistry and Biophysics . 742 109638. doi :10.1016/j.abb.2023.109638 . PMC 10251735 . PMID 37192692 . 1 2 3 4 Novák, Lukáš V. F.; Treitli, Sebastian C.; Pyrih, Jan; Hałakuc, Paweł; Pipaliya, Shweta V.; Vacek, Vojtěch; Brzoň, Ondřej; Soukal, Petr; Eme, Laura; Dacks, Joel B.; Karnkowska, Anna; Eliáš, Marek; Hampl, Vladimír (2023-12-07). Dutcher, Susan K. (ed.). "Genomics of Preaxostyla Flagellates Illuminates the Path Towards the Loss of Mitochondria" . PLOS Genetics . 19 (12) e1011050. doi : 10.1371/journal.pgen.1011050 . ISSN 1553-7404 . PMC 10703272 . PMID 38060519 . ↑ Anderson, Iain J.; Loftus, Brendan J. (2005). "Entamoeba histolytica: Observations on metabolism based on the genome sequence" . Experimental Parasitology . 110 (3): 173– 177. doi :10.1016/j.exppara.2005.03.010 . PMID 15955308 .
This page is based on this
Wikipedia article Text is available under the
CC BY-SA 4.0 license; additional terms may apply.
Images, videos and audio are available under their respective licenses.