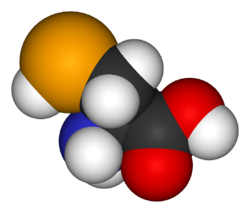
Selenocysteine is the Se-analogue of cysteine. It is rarely encountered outside of living tissue (nor is it available commercially) because of its high susceptiblility to air-oxidation. More common is the oxidized derivative selenocystine, which has an Se-Se bond.[9] Both selenocysteine and selenocystine are white solids. The Se-H group is more acidic (pKa = 5.43[3]) than the thiol group; thus, it is deprotonated at physiological pH.[10]
Structure
Selenocysteine has the same structure as cysteine, but with an atom of selenium taking the place of the usual sulfur; it has a selenol group. Like other natural proteinogenic amino acids, cysteine and selenocysteine have Lchirality in the older D/L notation based on homology to D- and L-glyceraldehyde. In the newer R/S system of designating chirality, based on the atomic numbers of atoms near the asymmetric carbon, they have R chirality, because of the presence of sulfur or selenium as a second neighbor to the asymmetric carbon. The remaining chiral amino acids, having only lighter atoms in that position, have S chirality.)
Proteins which contain a selenocysteine residue are called selenoproteins. Most selenoproteins contain a single selenocysteine residue. Selenoproteins that exhibit catalytic activity are called selenoenzymes.[11]
Biology
Unlike the other amino acids, no free pool of selenocysteine exists in the cell. Its high reactivity would cause damage to cells.[12] Instead, cells store selenium in the less reactive oxidized form, selenocystine, or in methylated form, selenomethionine.
Selenocysteine synthesis occurs on a specialized tRNA,[13] which also functions to incorporate it into nascent polypeptides. The primary and secondary structure of selenocysteine-specific tRNA, tRNASec, differ from those of standard tRNAs in several respects, most notably in having an 8-base-pair (bacteria) or 10-base-pair (eukaryotes)[Archaea?] acceptor stem, a long variable region arm, and substitutions at several well-conserved base positions. The selenocysteine tRNAs are initially charged with serine by seryl-tRNA ligase, but the resulting Ser-tRNASec is not used for translation because it is not recognised by the normal translation elongation factor (EF-Tu in bacteria, eEF1A in eukaryotes).[Archaea?]
Selenocysteine synthesis
Rather, the tRNA-bound seryl residue is converted to a selenocysteine residue by the pyridoxal phosphate-containing enzyme[14]selenocysteine synthase. In eukaryotes and archaea, two enzymes[15] are required to convert tRNA-bound seryl[16] residue into tRNA selenocysteinyl residue: PSTK (O-phosphoseryl-tRNA[Ser]Sec kinase) and selenocysteine synthase.[17][18] Finally, the resulting Sec-tRNASecis specifically bound to an alternative translational elongation factor (SelB[19] or mSelB (or eEFSec)), which delivers it in a targeted manner to the ribosomes[20] translating mRNAs[21][22] for selenoproteins. The specificity of this delivery mechanism is brought about by the presence of an extra protein domain (in bacteria, SelB) or an extra subunit (SBP2 for eukaryotic mSelB/eEFSec) which bind to the corresponding RNA secondary structures formed by the SECIS elements in selenoprotein mRNAs.
Selenoproteins
Selenocysteine has a lower reduction potential than cysteine. These properties make it very suitable in proteins that are involved in antioxidant activity.[23]
Although it is found in the three domains of life, it is not universal in all organisms.[24] Unlike other amino acids present in biological proteins, selenocysteine is not coded for directly in the genetic code.[25] Instead, it is encoded in a special way by a UGA codon, which is normally the "opal" stop codon. Such a mechanism is called translational recoding[26] and its efficiency depends on the selenoprotein being synthesized and on translation initiation factors.[27] When cells are grown in the absence of selenium, translation of selenoproteins terminates at the UGA codon, resulting in a truncated, nonfunctional enzyme. The UGA codon is made to encode selenocysteine by the presence of a selenocysteine insertion sequence (SECIS) in the mRNA. The SECIS element is defined by characteristic nucleotide sequences and secondary structure base-pairing patterns. In bacteria, the SECIS element is typically located immediately following the UGA codon within the reading frame for the selenoprotein.[28] In Archaea and in eukaryotes, the SECIS element is in the 3′ untranslated region (3′ UTR) of the mRNA and can direct multiple UGA codons to encode selenocysteine residues.[29]
As of 2021[update], 136 human proteins (in 37 families) are known to contain selenocysteine (selenoproteins).[30]
Breakdown
Selenocysteine is decomposed by the enzyme selenocysteine lyase into L-alanine and selenide. This probably helps with the safe recycling of Sec during degradation of selenoproteins.[31]
Toxicity
Just as selenomethionine can be randomly incorporated into proteins, selenocystine can also be mistakenly attached to tRNACys by cysteinyl-tRNA synthetase and incorporated into proteins in lieu of cystine. This causes considerable toxicity. A variant synthase that can distinguish between Cys and Sec helps reduce toxicity.[32]
Biotechnological applications of selenocysteine include use of 73Se-labeled Sec (half-life of 73Se = 7.2 hours) in positron emission tomography (PET) studies and 75Se-labeled Sec (half-life of 75Se = 118.5 days) in specific radiolabeling, facilitation of phase determination by multiwavelength anomalous diffraction in X-ray crystallography of proteins by introducing Sec alone, or Sec together with selenomethionine (SeMet), and incorporation of the stable 77Se isotope, which has a nuclear spin of 1/2 and can be used for high-resolution NMR, among others.[6]
See also
Pyrrolysine, another amino acid not in the basic set of 20.
Selenomethionine, another selenium-containing amino acid, which is randomly substituted for methionine.
↑"NIH 3D - Crystal structure of the selenocysteine synthase SelA and tRNASec complex." Nih.gov, 2023, https://3d.nih.gov/entries/2523. Accessed 5 Dec. 2025.
↑“NIH 3D - Human 80S Ribosome.” Nih.gov, 2019, 3d.nih.gov/entries/3DPX-011099. Accessed 5 Dec. 2025.
↑Longtin R (April 2004). "A forgotten debate: is selenocysteine the 21st amino acid?". Journal of the National Cancer Institute. 96 (7): 504–5. doi:10.1093/jnci/96.7.504. PMID15069108.
↑Böck A, Forchhammer K, Heider J, Baron C (December 1991). "Selenoprotein synthesis: an expansion of the genetic code". Trends in Biochemical Sciences. 16 (12): 463–7. doi:10.1016/0968-0004(91)90180-4. PMID1838215.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.