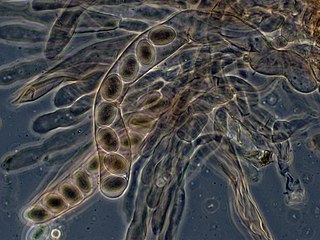
An ascus is the sexual spore-bearing cell produced in ascomycete fungi. Each ascus usually contains eight ascospores, produced by meiosis followed, in most species, by a mitotic cell division. However, asci in some genera or species can occur in numbers of one, two, four, or multiples of four. In a few cases, the ascospores can bud off conidia that may fill the asci with hundreds of conidia, or the ascospores may fragment, e.g. some Cordyceps, also filling the asci with smaller cells. Ascospores are nonmotile, usually single celled, but not infrequently may be coenocytic, and in some cases coenocytic in multiple planes. Mitotic divisions within the developing spores populate each resulting cell in septate ascospores with nuclei. The term ocular chamber, or oculus, refers to the epiplasm that is surrounded by the "bourrelet".

The Pertusariales are an order of fungi in the class Lecanoromycetes, comprising 8 families, 31 genera, and over 600 species, many of which form lichens. This diverse group is characterized by complex taxonomic history and ongoing phylogenetic revisions. Originally proposed by Maurice Choisy in 1949 and later formally published by the lichenologists David L. Hawksworth and Ove Eriksson in 1986, Pertusariales has undergone significant reclassification due to molecular phylogenetics studies. The order includes well-known genera such as Pertusaria and Ochrolechia, as well as families like Megasporaceae and Icmadophilaceae.

Japewia is a genus of lichen-forming fungi in the family Lecanoraceae. The genus was circumscribed in 1990 by the Norwegian lichenologist Tor Tønsberg, who assigned J. tornoënsis as the type species. The new genus is named in honour of Peter Wilfred James, a notable lichenologist, with Japewia being derived from the first letters of his initials (Ja) and his surname (Pe) followed by a typical Latin suffix (-wia).

Badimiella is a genus of lichen-forming fungi in the family Pilocarpaceae. It has two species of foliicolous (leaf-dwelling) lichens.

Mazosia is a genus of lichen-forming fungi in the family Roccellaceae.
Podotara is a fungal genus in the family Pilocarpaceae. It is a monotypic genus, containing the single species Podotara pilophoriformis, an uncommon foliicolous (leaf-dwelling), crustose lichen that grows on Podocarpus totara, a species of podocarp tree endemic to New Zealand. Both the genus and the species were proposed in 1996.
Fauriea is a genus of lichen-forming fungi in the family Teloschistaceae. The genus, which contains seven species, is a member of the subfamily Caloplacoideae.
Caloplaca letrouitioides is a little-known species of corticolous (bark-dwelling), crustose lichen belonging to the family Teloschistaceae, described in 2011. It is known to occur in Victoria, Australia. The species was named for its superficial resemblance to species in the genus Letrouitia. The anatomical characteristics of Caloplaca letrouitioides, particularly the well-developed true exciple and the unexpanded paraphyses tips, along with the absence of algae in the apothecia, set it apart from other species in the genus.
Megalospora austropacifica is a species of corticolous (bark-dwelling), crustose lichen in the family Megalosporaceae. It is found on the islands of Taveuni and Viti Levu in Fiji. It has a yellowish grey to whitish grey, glossy thallus that is thick and may appear slightly wrinkled or smooth, often with irregular cracks and small papillae containing conidiomata, but lacking isidia and soredia. Its apothecia are circular, up to 4.5 mm in diameter, with the disc evolving from concave to slightly convex and coloured from orange-brown to red-brown, surrounded by a thick, prominent margin.

Chaenothecopsis kilimanjaroensis is a species of lichenicolous (lichen-dwelling) pin lichen in the family Mycocaliciaceae. Found in the cloud forests of Tanzania, it was described as a new species in 2019. These tiny lichens have a short stalk, which can be either single or formed in aggregates on the same thallus. The stalks are medium brown at the base and become translucent in water. This species has unique spores, which contain a single septum, are arranged in a single row in the ascus, and have a surface ornamented with elongated, blister-like structures.
Harusavskia is single-species fungal genus in the family Teloschistaceae. It contains the little-known species Harusavskia elenkinianoides, a saxicolous (rock-dwelling), crustose lichen. This species is known only from its original collection site near the Laguna del Maule in Chile.

Glaucomaria is a genus of lichen-forming fungi in the family Lecanoraceae. It has seven species. The genus was circumscribed by Maurice Choisy in 1929. It contains crustose lichens formerly placed in the Lecanora rupicola species complex as defined by several previous authors.
Tapellaria intermedia is a little-known species of foliicolous (leaf-dwelling) crustose lichen in the family Pilocarpaceae. It occurs in Bolivia.
Synarthothelium is a genus of lichen-forming fungi of uncertain familial placement in the order Arthoniales. It has two species of corticolous (bark-dwelling) crustose lichens that occur in tropical regions of the Americas.

Gloeoheppia is a genus of lichen-forming fungi in the family Gloeoheppiaceae. It comprises five species. The genus is distinguished from similar-looking lichens like Heppia by its internal structure, the nature of its photobiont, and details of its reproductive structures.
Joergensenia is a fungal genus in the family Pannariaceae. It comprises a single species, Joergensenia cephalodina, which is a corticolous (bark-dwelling), squamulose lichen found in southern South America.
Megaloblastenia is a genus of crustose lichen-forming fungi in the family Megalosporaceae, comprising three species. Proposed by Dutch lichenologist Harrie Sipman in 1983, the genus is characterised by its thick, ecorticate thallus ranging from pale whitish-grey to yellowish, and its disc-like fruiting bodies (apothecia) that can be biatorine or lecideine. Megaloblastenia lichens form a symbiotic relationship with Dictyochloropsis algae, produce hyaline, bicellular ascospores with polaribilocular structure, and contain chemical compounds such as zeorin, pannarin, or usnic acid. Found in Australasia and South America, these lichens typically grow as epiphytes on trees in moist forests within temperate to tropical oceanic climates.

Paracollema is a small genus of lichen-forming fungi in the family Collemataceae. It comprises two species of jelly lichens, characterised by their small size, gelatinous nature when wet, and distinctive reproductive structures. The genus was proposed in 2013 and later validated in 2017. Paracollema lichens form small, leafy thalli up to 1 cm in diameter, with dark olive green to brownish colouration. They are distinguished from related genera by their very small asci and spores. Both known species are primarily epiphytic and have a limited distribution in southern Europe and northern Africa, typically found in Mediterranean or semi-arid climates.
Tylocliostomum is a fungal genus in the family Ramalinaceae. It comprises the single species Tylocliostomum viridifarinosum.