
Hyoliths are animals with small conical shells, known as fossils from the Palaeozoic era. They are at least considered as lophotrochozoan, and possibly being lophophorates, a group which includes the brachiopods, while others consider them as being basal lophotrochozoans, or even molluscs.

The tusk shells or tooth shells, technically the Scaphopoda, are members of a class of shelled marine mollusc with worldwide distribution, and are the only class of exclusively infaunal marine molluscs. Shells of species within this class range in length 0.5–18 cm (0.20–7.09 in). Members of the order Dentaliida tend to be larger than those of the order Gadilida.

The Rostroconchia is a class of extinct molluscs dating from the early Cambrian to the Late Permian. They were initially thought to be bivalves, but were later given their own class. They have a single shell in their larval stage, and the adult typically has a single, pseudo-bivalved shell enclosing the mantle and muscular foot. The anterior part of the shell probably pointed downward and had a gap from which the foot could probably emerge. Rostroconchs probably lived a sedentary semi-infaunal lifestyle. There were probably more than 1,000 species of members of this class.
The Coelosclerithophorans are a polyphyletic group of organisms bearing hollow sclerites made of aragonite, and with a supposedly distinctive microstructure.
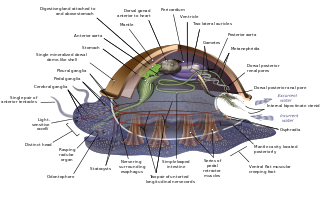
The evolution of the molluscs is the way in which the Mollusca, one of the largest groups of invertebrate animals, evolved. This phylum includes gastropods, bivalves, scaphopods, cephalopods, and several other groups. The fossil record of mollusks is relatively complete, and they are well represented in most fossil-bearing marine strata. Very early organisms which have dubiously been compared to molluscs include Kimberella and Odontogriphus.

Yochelcionella is an extinct genus of basal molluscs which lived during the Tommotian epoch, the first epoch of the Cambrian period. This genus is often reconstructed to resemble snails.

Helcionellid or Helcionelliformes is an order of small fossil shells that are universally interpreted as molluscs, though no sources spell out why this taxonomic interpretation is preferred. These animals are first found about 540 to 530 million years ago in the late Nemakit-Daldynian age, which is the earliest part of the Cambrian period. A single species persisted to the Early Ordovician. These fossils are component of the small shelly fossils (SSF) assemblages.
The small shelly fauna, small shelly fossils (SSF), or early skeletal fossils (ESF) are mineralized fossils, many only a few millimetres long, with a nearly continuous record from the latest stages of the Ediacaran to the end of the Early Cambrian Period. They are very diverse, and there is no formal definition of "small shelly fauna" or "small shelly fossils". Almost all are from earlier rocks than more familiar fossils such as trilobites. Since most SSFs were preserved by being covered quickly with phosphate and this method of preservation is mainly limited to the late Ediacaran and early Cambrian periods, the animals that made them may actually have arisen earlier and persisted after this time span.

Mollusca is the second-largest phylum of invertebrate animals, after Arthropoda; members are known as molluscs or mollusks. Around 76,000 extant species of molluscs are recognized. The number of fossil species is estimated between 60,000 and 100,000 additional species. The proportion of undescribed species is very high. Many taxa remain poorly studied.
Stenothecidae is an extinct family of fossil univalved Cambrian molluscs which may be either gastropods or monoplacophorans.
Anabarella is a species of bilaterally-flattened monoplacophoran mollusc, with a morphological similarity to the rostroconchs. Its shell preserves evidence of three mineralogical textures on its outer surface: it is polygonal near the crest of the shell, subsequently changing to both spiny and stepwise. Its internal microstructure is calcitic and semi-nacreous. Its name reflects its provenance from Anabar, Siberia. It has been interpreted as ancestral to the rostroconchs, and has been aligned to the Helcionellidae.

Fordilla is an extinct genus of early bivalves, one of two genera in the extinct family Fordillidae. The genus is known solely from Early Cambrian fossils found in North America, Greenland, Europe, the Middle East, and Asia. The genus currently contains three described species, Fordilla germanica, Fordilla sibirica, and the type species Fordilla troyensis.
Stenothecoida is a taxon of bivalved fossils from the Early to middle Cambrian period. They look a bit like brachiopods or bivalve molluscs.
Ribeiria is a genus of rostroconch mollusc with a pegma, once interpreted as an intermediate between helcionellids and primitive scaphopods.
Pojetaia is an extinct genus of early bivalves, one of two genera in the extinct family Fordillidae. The genus is known solely from Early to Middle Cambrian fossils found in North America, Greenland, Europe, North Africa, Asia, and Australia. The genus currently contains two accepted species, Pojetaia runnegari, the type species, and Pojetaia sarhroensis, though up to seven species have been proposed. The genera Buluniella, Jellia, and Oryzoconcha are all considered synonyms of Pojetaia.

Fordillidae is an extinct family of early bivalves and one of two families in the extinct superfamily Fordilloidea. The family is known from fossils of early to middle Cambrian age found in North America, Greenland, Europe, the Middle East, Asia, and Australia. The family currently contains two genera, Fordilla and Pojetaia, each with up to three described species. Due to the size and age of the fossil specimens, Fordillidae species are included as part of the Turkish Small shelly fauna.
Fordilloidea is an extinct superfamily of early bivalves containing two described families, Fordillidae and Camyidae and the only superfamily in the order Fordillida. The superfamily is known from fossils of early to middle Cambrian age found in North America, Greenland, Europe, the Middle East, Asia, and Australia. Fordillidae currently contains two genera, Fordilla and Pojetaia each with up to three described species while Camyidae only contains a single genus Camya with one described species, Camya asy. Due to the size and age of the fossil specimens, Fordillidae species are included as part of the Turkish Small shelly fauna.
Tuarangia is a Cambrian shelly fossil interpreted as an early bivalve, though alternative classifications have been proposed and its systematic position remains controversial. It is the only genus in the extinct family Tuarangiidae and order Tuarangiida. The genus is known solely from Middle to Late Cambrian fossils found in Europe and New Zealand. The genus currently contains two accepted species, Tuarangia gravgaerdensis and the type species Tuarangia paparua.
Stage 2 of the Cambrian is the unnamed upper stage of the Terreneuvian Series. It lies atop the Fortunian and below Stage 3 of the Cambrian. It is commonly referred to as the Tommotian, after the Cambrian stratigraphy of Siberia. Neither the upper nor lower boundary has yet been defined by the International Commission on Stratigraphy. The preferred definitions for the lower boundary are the first appearance of the molluscs Watsonella crosbyi or Aldanella attleborensis around 529 million years ago. The correlation between the lowest occurrences of A. attleborensis and W. crosbyi with the Zhujiaqing positive carbon isotope excursion (ZHUCE) has led to the conclusion that the combination of these two markers are a reliable indicator of the boundary between the Fortunian and Stage 2. The proposed upper boundary might be the first appearance of trilobites around 521 million years ago.

Bruce Norman Runnegar is an Australian-born paleontologist and professor at UCLA. His research centers on using the fossil record to determine how, where, and when life originated and evolved. He has published on a wide variety of topics, including the phylogeny of molluscs, Dickinsonia fossils and oxygen levels, and molecular clock techniques.