
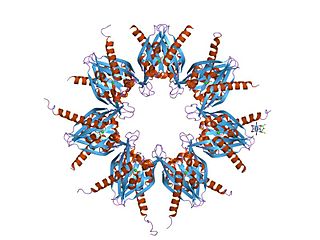
The α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptor (also known as AMPA receptor, AMPAR, or quisqualate receptor) is an ionotropic transmembrane receptor for glutamate (iGluR) and predominantly Na+ ion channel that mediates fast synaptic transmission in the central nervous system (CNS). It has been traditionally classified as a non-NMDA-type receptor, along with the kainate receptor. Its name is derived from its ability to be activated by the artificial glutamate analog AMPA. The receptor was first named the "quisqualate receptor" by Watkins and colleagues after a naturally occurring agonist quisqualate and was only later given the label "AMPA receptor" after the selective agonist developed by Tage Honore and colleagues at the Royal Danish School of Pharmacy in Copenhagen. The GRIA2-encoded AMPA receptor ligand binding core (GluA2 LBD) was the first glutamate receptor ion channel domain to be crystallized.

The N-methyl-D-aspartatereceptor (also known as the NMDA receptor or NMDAR), is a glutamate receptor and predominantly Ca2+ ion channel found in neurons. The NMDA receptor is one of three types of ionotropic glutamate receptors, the other two being AMPA and kainate receptors. Depending on its subunit composition, its ligands are glutamate and glycine (or D-serine). However, the binding of the ligands is typically not sufficient to open the channel as it may be blocked by Mg2+ ions which are only removed when the neuron is sufficiently depolarized. Thus, the channel acts as a "coincidence detector" and only once both of these conditions are met, the channel opens and it allows positively charged ions (cations) to flow through the cell membrane. The NMDA receptor is thought to be very important for controlling synaptic plasticity and mediating learning and memory functions.

Brain-derived neurotrophic factor (BDNF), or abrineurin, is a protein that, in humans, is encoded by the BDNF gene. BDNF is a member of the neurotrophin family of growth factors, which are related to the canonical nerve growth factor (NGF), a family which also includes NT-3 and NT-4/NT-5. Neurotrophic factors are found in the brain and the periphery. BDNF was first isolated from a pig brain in 1982 by Yves-Alain Barde and Hans Thoenen.
Immediate early genes (IEGs) are genes which are activated transiently and rapidly in response to a wide variety of cellular stimuli. They represent a standing response mechanism that is activated at the transcription level in the first round of response to stimuli, before any new proteins are synthesized. IEGs are distinct from "late response" genes, which can only be activated later, following the synthesis of early response gene products. Thus IEGs have been called the "gateway to the genomic response". The term can describe viral regulatory proteins that are synthesized following viral infection of a host cell, or cellular proteins that are made immediately following stimulation of a resting cell by extracellular signals.
Synaptogenesis is the formation of synapses between neurons in the nervous system. Although it occurs throughout a healthy person's lifespan, an explosion of synapse formation occurs during early brain development, known as exuberant synaptogenesis. Synaptogenesis is particularly important during an individual's critical period, during which there is a certain degree of synaptic pruning due to competition for neural growth factors by neurons and synapses. Processes that are not used, or inhibited during their critical period will fail to develop normally later on in life.
Neuromodulation is the physiological process by which a given neuron uses one or more chemicals to regulate diverse populations of neurons. Neuromodulators typically bind to metabotropic, G-protein coupled receptors (GPCRs) to initiate a second messenger signaling cascade that induces a broad, long-lasting signal. This modulation can last for hundreds of milliseconds to several minutes. Some of the effects of neuromodulators include: altering intrinsic firing activity, increasing or decreasing voltage-dependent currents, altering synaptic efficacy, increasing bursting activity and reconfigurating synaptic connectivity.

The p75 neurotrophin receptor (p75NTR) was first identified in 1973 as the low-affinity nerve growth factor receptor (LNGFR) before discovery that p75NTR bound other neurotrophins equally well as nerve growth factor. p75NTR is a neurotrophic factor receptor. Neurotrophic factor receptors bind Neurotrophins including Nerve growth factor, Neurotrophin-3, Brain-derived neurotrophic factor, and Neurotrophin-4. All neurotrophins bind to p75NTR. This also includes the immature pro-neurotrophin forms. Neurotrophic factor receptors, including p75NTR, are responsible for ensuring a proper density to target ratio of developing neurons, refining broader maps in development into precise connections. p75NTR is involved in pathways that promote neuronal survival and neuronal death.
Ca2+
/calmodulin-dependent protein kinase II is a serine/threonine-specific protein kinase that is regulated by the Ca2+
/calmodulin complex. CaMKII is involved in many signaling cascades and is thought to be an important mediator of learning and memory. CaMKII is also necessary for Ca2+
homeostasis and reuptake in cardiomyocytes, chloride transport in epithelia, positive T-cell selection, and CD8 T-cell activation.

Kalirin, also known as Huntingtin-associated protein-interacting protein (HAPIP), protein duo (DUO), or serine/threonine-protein kinase with Dbl- and pleckstrin homology domain, is a protein that in humans is encoded by the KALRN gene. Kalirin was first identified in 1997 as a protein interacting with huntingtin-associated protein 1. Is also known to play an important role in nerve growth and axonal development.

Glutamate receptor 1 is a protein that in humans is encoded by the GRIA1 gene.

Protein phosphatase 1 regulatory subunit 1B (PPP1R1B), also known as dopamine- and cAMP-regulated neuronal phosphoprotein (DARPP-32), is a protein that in humans is encoded by the PPP1R1B gene.

Homer protein homolog 1 or Homer1 is a neuronal protein that in humans is encoded by the HOMER1 gene. Other names are Vesl and PSD-Zip45.

POU domain, class 4, transcription factor 1 (POU4F1) also known as brain-specific homeobox/POU domain protein 3A (BRN3A), homeobox/POU domain protein RDC-1 or Oct-T1 is a protein that in humans is encoded by the POU4F1 gene.

Synaptic Ras GTPase-activating protein 1, also known as synaptic Ras-GAP 1 or SYNGAP1, is a protein that in humans is encoded by the SYNGAP1 gene. SYNGAP1 is a ras GTPase-activating protein that is critical for the development of cognition and proper synapse function. Mutations in humans can cause intellectual disability, epilepsy, autism and sensory processing deficits.

Potassium-chloride transporter member 5 is a neuron-specific chloride potassium symporter responsible for establishing the chloride ion gradient in neurons through the maintenance of low intracellular chloride concentrations. It is a critical mediator of synaptic inhibition, cellular protection against excitotoxicity and may also act as a modulator of neuroplasticity. Potassium-chloride transporter member 5 is also known by the names: KCC2 for its ionic substrates, and SLC12A5 for its genetic origin from the SLC12A5 gene in humans.

Cyclin-dependent kinase 5 is a protein, and more specifically an enzyme, that is encoded by the Cdk5 gene. It was discovered 15 years ago, and it is saliently expressed in post-mitotic central nervous system neurons (CNS).
Activity-dependent plasticity is a form of functional and structural neuroplasticity that arises from the use of cognitive functions and personal experience; hence, it is the biological basis for learning and the formation of new memories. Activity-dependent plasticity is a form of neuroplasticity that arises from intrinsic or endogenous activity, as opposed to forms of neuroplasticity that arise from extrinsic or exogenous factors, such as electrical brain stimulation- or drug-induced neuroplasticity. The brain's ability to remodel itself forms the basis of the brain's capacity to retain memories, improve motor function, and enhance comprehension and speech amongst other things. It is this trait to retain and form memories that is associated with neural plasticity and therefore many of the functions individuals perform on a daily basis. This plasticity occurs as a result of changes in gene expression which are triggered by signaling cascades that are activated by various signaling molecules during increased neuronal activity.
While the cellular and molecular mechanisms of learning and memory have long been a central focus of neuroscience, it is only in recent years that attention has turned to the epigenetic mechanisms behind the dynamic changes in gene transcription responsible for memory formation and maintenance. Epigenetic gene regulation often involves the physical marking of DNA or associated proteins to cause or allow long-lasting changes in gene activity. Epigenetic mechanisms such as DNA methylation and histone modifications have been shown to play an important role in learning and memory.
Intermediate-term memory (ITM) is a stage of memory distinct from sensory memory, working memory/short-term memory, and long-term memory. While sensory memory persists for several milliseconds, working memory persists for up to thirty seconds, and long-term memory persists from thirty minutes to the end of an individual's life, intermediate-term memory persists for about two to three hours. This overlap in the durations of these memory processes indicates that they occur simultaneously, rather than sequentially. Indeed, intermediate-term facilitation can be produced in the absence of long-term facilitation. However, the boundaries between these forms of memory are not clear-cut, and they can vary depending on the task. Intermediate-term memory is thought to be supported by the parahippocampal cortex.
R. Suzanne Zukin is an American neuroscientist and a professor of neuroscience who directs a research lab as a F. M. Kirby Chair in Neural Repair and Protection and director of the Neuropsychopharmacology Center at Albert Einstein College of Medicine. Zukin's areas of research include neurodegenerative disorders, Ischemia, Epigenetics and Autism and uses molecular biology approaches to research these disorders. Zukin has made seminal contributions to the understanding of NMDA and AMPA receptor function and activity.



