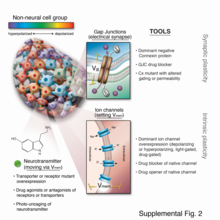
Ion channels are pore-forming membrane proteins that allow ions to pass through the channel pore. Their functions include establishing a resting membrane potential, shaping action potentials and other electrical signals by gating the flow of ions across the cell membrane, controlling the flow of ions across secretory and epithelial cells, and regulating cell volume. Ion channels are present in the membranes of all cells. Ion channels are one of the two classes of ionophoric proteins, the other being ion transporters.

In vertebrates, most neuronal cell axons are encased in myelin. Simply put, myelin insulates axons and increases the rate at which electrical impulses are passed along the axon. The myelinated axon can be likened to an electrical wire with insulating material (myelin) around it. However, unlike the plastic covering on an electrical wire, myelin does not form a single long sheath over the entire length of the axon. Rather, myelin ensheaths the axon in segments: in general, each axon is encased in multiple long myelin sheaths separated by short gaps called nodes of Ranvier.
Beta cells (β-cells) are a type of cell found in pancreatic islets that synthesize and secrete insulin and amylin. Beta cells make up 50–70% of the cells in human islets. In patients with Type 1 diabetes, beta-cell mass and function are diminished, leading to insufficient insulin secretion and hyperglycemia.
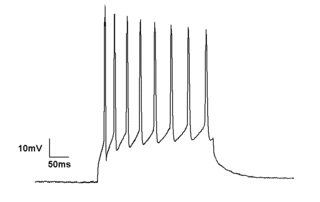
Electrophysiology is the branch of physiology that studies the electrical properties of biological cells and tissues. It involves measurements of voltage changes or electric current or manipulations on a wide variety of scales from single ion channel proteins to whole organs like the heart. In neuroscience, it includes measurements of the electrical activity of neurons, and, in particular, action potential activity. Recordings of large-scale electric signals from the nervous system, such as electroencephalography, may also be referred to as electrophysiological recordings. They are useful for electrodiagnosis and monitoring.

An action potential occurs when the membrane potential of a specific cell rapidly rises and falls. This depolarization then causes adjacent locations to similarly depolarize. Action potentials occur in several types of animal cells, called excitable cells, which include neurons, muscle cells, and in some plant cells. Certain endocrine cells such as pancreatic beta cells, and certain cells of the anterior pituitary gland are also excitable cells.

Electrophorus electricus is the best-known species of electric eel. It is a South American electric fish. Until the discovery of two additional species in 2019, the genus was classified as the monotypic, with this species the only one in the genus. Despite the name, it is not an eel, but rather a knifefish. It is considered as a freshwater teleost which contains an electrogenic tissue that produces electric discharges.

Membrane potential is the difference in electric potential between the interior and the exterior of a biological cell. That is, there is a difference in the energy required for electric charges to move from the internal to exterior cellular environments and vice versa, as long as there is no acquisition of kinetic energy or the production of radiation. The concentration gradients of the charges directly determine this energy requirement. For the exterior of the cell, typical values of membrane potential, normally given in units of milli volts and denoted as mV, range from –80 mV to –40 mV.
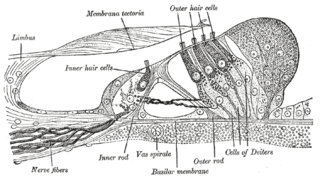
Hair cells are the sensory receptors of both the auditory system and the vestibular system in the ears of all vertebrates, and in the lateral line organ of fishes. Through mechanotransduction, hair cells detect movement in their environment.

An electric fish is any fish that can generate electric fields. Most electric fish are also electroreceptive, meaning that they can sense electric fields. The only exception is the stargazer family (Uranoscopidae). Electric fish, although a small minority of all fishes, include both oceanic and freshwater species, and both cartilaginous and bony fishes.
Voltage-gated calcium channels (VGCCs), also known as voltage-dependent calcium channels (VDCCs), are a group of voltage-gated ion channels found in the membrane of excitable cells (e.g., muscle, glial cells, neurons, etc.) with a permeability to the calcium ion Ca2+. These channels are slightly permeable to sodium ions, so they are also called Ca2+-Na+ channels, but their permeability to calcium is about 1000-fold greater than to sodium under normal physiological conditions.
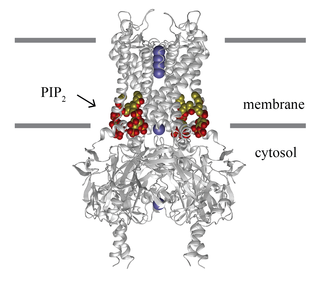
Inward-rectifier potassium channels (Kir, IRK) are a specific lipid-gated subset of potassium channels. To date, seven subfamilies have been identified in various mammalian cell types, plants, and bacteria. They are activated by phosphatidylinositol 4,5-bisphosphate (PIP2). The malfunction of the channels has been implicated in several diseases. IRK channels possess a pore domain, homologous to that of voltage-gated ion channels, and flanking transmembrane segments (TMSs). They may exist in the membrane as homo- or heterooligomers and each monomer possesses between 2 and 4 TMSs. In terms of function, these proteins transport potassium (K+), with a greater tendency for K+ uptake than K+ export. The process of inward-rectification was discovered by Denis Noble in cardiac muscle cells in 1960s and by Richard Adrian and Alan Hodgkin in 1970 in skeletal muscle cells.
Sodium channels are integral membrane proteins that form ion channels, conducting sodium ions (Na+) through a cell's membrane. They belong to the superfamily of cation channels.
Bioelectromagnetics, also known as bioelectromagnetism, is the study of the interaction between electromagnetic fields and biological entities. Areas of study include electromagnetic fields produced by living cells, tissues or organisms, the effects of man-made sources of electromagnetic fields like mobile phones, and the application of electromagnetic radiation toward therapies for the treatment of various conditions.

Calcium channel, voltage-dependent, L type, alpha 1C subunit is a protein that in humans is encoded by the CACNA1C gene. Cav1.2 is a subunit of L-type voltage-dependent calcium channel.

Voltage-dependent anion-selective channel protein 2 is a protein that in humans is encoded by the VDAC2 gene on chromosome 10. This protein is a voltage-dependent anion channel and shares high structural homology with the other VDAC isoforms. VDACs are generally involved in the regulation of cell metabolism, mitochondrial apoptosis, and spermatogenesis. Additionally, VDAC2 participates in cardiac contractions and pulmonary circulation, which implicate it in cardiopulmonary diseases. VDAC2 also mediates immune response to infectious bursal disease (IBD).
In electrophysiology, the term gating refers to the opening (activation) or closing of ion channels. This change in conformation is a response to changes in transmembrane voltage.
In biology, electrotropism, also known as galvanotropism, is a kind of tropism which results in growth or migration of an organism, usually a cell, in response to an exogenous electric field. Several types of cells such as nerve cells, muscle cells, fibroblasts, epithelial cells, green algae, spores, and pollen tubes, among others, have been already reported to respond by either growing or migrating in a preferential direction when exposed to an electric field.
Scanning vibrating electrode technique (SVET), also known as vibrating probe within the field of biology, is a scanning probe microscopy (SPM) technique which visualizes electrochemical processes at a sample. It was originally introduced in 1974 by Jaffe and Nuccitelli to investigate the electrical current densities near living cells. Starting in the 1980s Hugh Isaacs began to apply SVET to a number of different corrosion studies. SVET measures local current density distributions in the solution above the sample of interest, to map electrochemical processes in situ as they occur. It utilizes a probe, vibrating perpendicular to the sample of interest, to enhance the measured signal. It is related to scanning ion-selective electrode technique (SIET), which can be used with SVET in corrosion studies, and scanning reference electrode technique (SRET), which is a precursor to SVET.
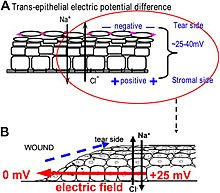
Electrotaxis, also known as galvanotaxis, is the directed motion of biological cells or organisms guided by an electric field or current. The directed motion of electrotaxis can take many forms, such as; growth, development, active swimming, and passive migration. A wide variety of biological cells can naturally sense and follow DC electric fields. Such electric fields arise naturally in biological tissues during development and healing. These and other observations have led to research into how applied electric fields can impact wound healing An increase in wound healing rate is regularly observed and this is thought to be due to the cell migration and other signaling pathways that are activated by the electric field. Additional research has been conducted into how applied electric fields impact cancer metastasis, morphogenesis, neuron guidance, motility of pathogenic bacteria, biofilm formation, and many other biological phenomena.

Michael Levin is an American developmental and synthetic biologist at Tufts University, where he is the Vannevar Bush Distinguished Professor. Levin is a director of the Allen Discovery Center at Tufts University and Tufts Center for Regenerative and Developmental Biology. He is also co-director of the Institute for Computationally Designed Organisms with Josh Bongard.