Related Research Articles

The visual cortex of the brain is the area of the cerebral cortex that processes visual information. It is located in the occipital lobe. Sensory input originating from the eyes travels through the lateral geniculate nucleus in the thalamus and then reaches the visual cortex. The area of the visual cortex that receives the sensory input from the lateral geniculate nucleus is the primary visual cortex, also known as visual area 1 (V1), Brodmann area 17, or the striate cortex. The extrastriate areas consist of visual areas 2, 3, 4, and 5.
Blindsight is the ability of people who are cortically blind to respond to visual stimuli that they do not consciously see due to lesions in the primary visual cortex, also known as the striate cortex or Brodmann Area 17. The term was coined by Lawrence Weiskrantz and his colleagues in a paper published in a 1974 issue of Brain. A previous paper studying the discriminatory capacity of a cortically blind patient was published in Nature in 1973. The assumed existence of blindsight is controversial, with some arguing that it is merely degraded conscious vision.

Color vision, a feature of visual perception, is an ability to perceive differences between light composed of different frequencies independently of light intensity.

The visual system is the physiological basis of visual perception. The system detects, transduces and interprets information concerning light within the visible range to construct an image and build a mental model of the surrounding environment. The visual system is associated with the eye and functionally divided into the optical system and the neural system.
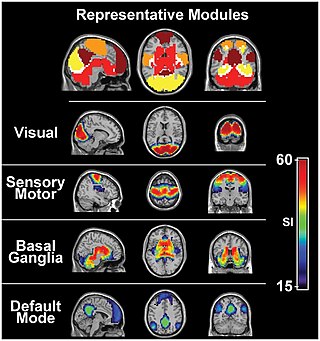
The sensory nervous system is a part of the nervous system responsible for processing sensory information. A sensory system consists of sensory neurons, neural pathways, and parts of the brain involved in sensory perception and interoception. Commonly recognized sensory systems are those for vision, hearing, touch, taste, smell, balance and visceral sensation. Sense organs are transducers that convert data from the outer physical world to the realm of the mind where people interpret the information, creating their perception of the world around them.

In neuroanatomy, the lateral geniculate nucleus is a structure in the thalamus and a key component of the mammalian visual pathway. It is a small, ovoid, ventral projection of the thalamus where the thalamus connects with the optic nerve. There are two LGNs, one on the left and another on the right side of the thalamus. In humans, both LGNs have six layers of neurons alternating with optic fibers.

The occipital lobe is one of the four major lobes of the cerebral cortex in the brain of mammals. The name derives from its position at the back of the head, from the Latin ob, 'behind', and caput, 'head'.

Brodmann area 19, or BA 19, is part of the occipital lobe cortex in the human brain. Along with area 18, it comprises the extrastriate cortex. In humans with normal sight, extrastriate cortex is a visual association area, with feature-extracting, shape recognition, attentional, and multimodal integrating functions.

A retinal ganglion cell (RGC) is a type of neuron located near the inner surface of the retina of the eye. It receives visual information from photoreceptors via two intermediate neuron types: bipolar cells and retina amacrine cells. Retina amacrine cells, particularly narrow field cells, are important for creating functional subunits within the ganglion cell layer and making it so that ganglion cells can observe a small dot moving a small distance. Retinal ganglion cells collectively transmit image-forming and non-image forming visual information from the retina in the form of action potential to several regions in the thalamus, hypothalamus, and mesencephalon, or midbrain.
In neuroscience, parvocellular cells, also called P-cells, are neurons located within the parvocellular layers of the lateral geniculate nucleus (LGN) of the thalamus. Their name comes from Latin parvus 'small', due to the small size of the cell compared to the larger magnocellular cells. Phylogenetically, parvocellular neurons are more modern than magnocellular ones.
Magnocellular cells, also called M-cells, are neurons located within the magnocellular layer of the lateral geniculate nucleus of the thalamus. The cells are part of the visual system. They are termed "magnocellular" since they are characterized by their relatively large size compared to parvocellular cells.
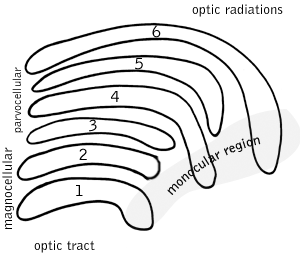
In neuroscience, koniocellular cells, also called K-cells, are relatively small neurons located in the koniocellular layer of the lateral geniculate nucleus (LGN) within the thalamus of primates, including humans. The term 'koniocellular' is derived from Greek konio 'dust, poison'.
Ocular dominance columns are stripes of neurons in the visual cortex of certain mammals that respond preferentially to input from one eye or the other. The columns span multiple cortical layers, and are laid out in a striped pattern across the surface of the striate cortex (V1). The stripes lie perpendicular to the orientation columns.

The inferior temporal gyrus is one of three gyri of the temporal lobe and is located below the middle temporal gyrus, connected behind with the inferior occipital gyrus; it also extends around the infero-lateral border on to the inferior surface of the temporal lobe, where it is limited by the inferior sulcus. This region is one of the higher levels of the ventral stream of visual processing, associated with the representation of objects, places, faces, and colors. It may also be involved in face perception, and in the recognition of numbers and words.

The colour centre is a region in the brain primarily responsible for visual perception and cortical processing of colour signals received by the eye, which ultimately results in colour vision. The colour centre in humans is thought to be located in the ventral occipital lobe as part of the visual system, in addition to other areas responsible for recognizing and processing specific visual stimuli, such as faces, words, and objects. Many functional magnetic resonance imaging (fMRI) studies in both humans and macaque monkeys have shown colour stimuli to activate multiple areas in the brain, including the fusiform gyrus and the lingual gyrus. These areas, as well as others identified as having a role in colour vision processing, are collectively labelled visual area 4 (V4). The exact mechanisms, location, and function of V4 are still being investigated.
Globs are millimeter-sized color modules found beyond the visual area V2 in the brain's color processing ventral pathway. They are scattered throughout the posterior inferior temporal cortex in an area called the V4 complex. They are clustered by color preference, and organized as color columns. They are the first part of the brain in which color is processed in terms of the full range of hues found in color space.

A parasol cell, sometimes called an M cell or M ganglion cell, is one type of retinal ganglion cell (RGC) located in the ganglion cell layer of the retina. These cells project to magnocellular cells in the lateral geniculate nucleus (LGN) as part of the magnocellular pathway in the visual system. They have large cell bodies as well as extensive branching dendrite networks and as such have large receptive fields. Relative to other RGCs, they have fast conduction velocities. While they do show clear center-surround antagonism, they receive no information about color. Parasol ganglion cells contribute information about the motion and depth of objects to the visual system.

Orientation columns are organized regions of neurons that are excited by visual line stimuli of varying angles. These columns are located in the primary visual cortex (V1) and span multiple cortical layers. The geometry of the orientation columns are arranged in slabs that are perpendicular to the surface of the primary visual cortex.
Binocular neurons are neurons in the visual system that assist in the creation of stereopsis from binocular disparity. They have been found in the primary visual cortex where the initial stage of binocular convergence begins. Binocular neurons receive inputs from both the right and left eyes and integrate the signals together to create a perception of depth.

Laura Busse is a German neuroscientist and professor of Systemic Neuroscience within the Division of Neurobiology at the Ludwig Maximilian University of Munich. Busse's lab studies context-dependent visual processing in mouse models by performing large scale in vivo electrophysiological recordings in the thalamic and cortical circuits of awake and behaving mice.
References
- ↑ Wong-Riley, Margaret (July 27, 1979). "Changes in the visual system of monocularly sutured or enucleated cats demonstrable with cytochrome oxidase histochemistry". Brain Research . 171 (1): 11–28. doi:10.1016/0006-8993(79)90728-5. PMID 223730. S2CID 20990578.
- ↑ Rockland, Kathleen S. (2021). "Cytochrome oxidase "blobs": a call for more anatomy". Brain Structure and Function. 226 (9): 2793–2806. doi:10.1007/s00429-021-02360-2. PMC 8778949 . PMID 34382115.
| Illusions |
| |
|---|---|---|
| Popular culture |
| |
| Related | ||
| | This neuroanatomy article is a stub. You can help Wikipedia by expanding it. |