
A motor neuron is a neuron whose cell body is located in the motor cortex, brainstem or the spinal cord, and whose axon (fiber) projects to the spinal cord or outside of the spinal cord to directly or indirectly control effector organs, mainly muscles and glands. There are two types of motor neuron – upper motor neurons and lower motor neurons. Axons from upper motor neurons synapse onto interneurons in the spinal cord and occasionally directly onto lower motor neurons. The axons from the lower motor neurons are efferent nerve fibers that carry signals from the spinal cord to the effectors. Types of lower motor neurons are alpha motor neurons, beta motor neurons, and gamma motor neurons.
In biology, a reflex, or reflex action, is an involuntary, unplanned sequence or action and nearly instantaneous response to a stimulus.

The somatic nervous system (SNS) is made up of nerves that link the brain and spinal cord to voluntary or skeletal muscles that are under conscious control as well as to skin sensory receptors. Specialized nerve fiber ends called sensory receptors are responsible for detecting information within and outside of the body.
Motor learning refers broadly to changes in an organism's movements that reflect changes in the structure and function of the nervous system. Motor learning occurs over varying timescales and degrees of complexity: humans learn to walk or talk over the course of years, but continue to adjust to changes in height, weight, strength etc. over their lifetimes. Motor learning enables animals to gain new skills, and improves the smoothness and accuracy of movements, in some cases by calibrating simple movements like reflexes. Motor learning research often considers variables that contribute to motor program formation, sensitivity of error-detection processes, and strength of movement schemas. Motor learning is "relatively permanent", as the capability to respond appropriately is acquired and retained. Temporary gains in performance during practice or in response to some perturbation are often termed motor adaptation, a transient form of learning. Neuroscience research on motor learning is concerned with which parts of the brain and spinal cord represent movements and motor programs and how the nervous system processes feedback to change the connectivity and synaptic strengths. At the behavioral level, research focuses on the design and effect of the main components driving motor learning, i.e. the structure of practice and the feedback. The timing and organization of practice can influence information retention, e.g. how tasks can be subdivided and practiced, and the precise form of feedback can influence preparation, anticipation, and guidance of movement.

Muscle spindles are stretch receptors within the body of a skeletal muscle that primarily detect changes in the length of the muscle. They convey length information to the central nervous system via afferent nerve fibers. This information can be processed by the brain as proprioception. The responses of muscle spindles to changes in length also play an important role in regulating the contraction of muscles, for example, by activating motor neurons via the stretch reflex to resist muscle stretch.
Neural engineering is a discipline within biomedical engineering that uses engineering techniques to understand, repair, replace, or enhance neural systems. Neural engineers are uniquely qualified to solve design problems at the interface of living neural tissue and non-living constructs.
Central pattern generators (CPGs) are self-organizing biological neural circuits that produce rhythmic outputs in the absence of rhythmic input. They are the source of the tightly-coupled patterns of neural activity that drive rhythmic and stereotyped motor behaviors like walking, swimming, breathing, or chewing. The ability to function without input from higher brain areas still requires modulatory inputs, and their outputs are not fixed. Flexibility in response to sensory input is a fundamental quality of CPG-driven behavior. To be classified as a rhythmic generator, a CPG requires:
- "two or more processes that interact such that each process sequentially increases and decreases, and
- that, as a result of this interaction, the system repeatedly returns to its starting condition."
Motor control is the regulation of movements in organisms that possess a nervous system. Motor control includes conscious voluntary movements, subconscious muscle memory and involuntary reflexes, as well as instinctual taxis.
In physiology, motor coordination is the orchestrated movement of multiple body parts as required to accomplish intended actions, like walking. This coordination is achieved by adjusting kinematic and kinetic parameters associated with each body part involved in the intended movement. The modifications of these parameters typically relies on sensory feedback from one or more sensory modalities, such as proprioception and vision.

The scratch reflex is a response to activation of sensory neurons whose peripheral terminals are located on the surface of the body. Some sensory neurons can be activated by stimulation with an external object such as a parasite on the body surface. Alternatively, some sensory neurons can respond to a chemical stimulus that produces an itch sensation. During a scratch reflex, a nearby limb reaches toward and rubs against the site on the body surface that has been stimulated. The scratch reflex has been extensively studied to understand the functioning of neural networks in vertebrates. Despite decades of research, key aspects of the scratch reflex are still unknown, such as the neural mechanisms by which the reflex is terminated.

The vestibulospinal tract is a neural tract in the central nervous system. Specifically, it is a component of the extrapyramidal system and is classified as a component of the medial pathway. Like other descending motor pathways, the vestibulospinal fibers of the tract relay information from nuclei to motor neurons. The vestibular nuclei receive information through the vestibulocochlear nerve about changes in the orientation of the head. The nuclei relay motor commands through the vestibulospinal tract. The function of these motor commands is to alter muscle tone, extend, and change the position of the limbs and head with the goal of supporting posture and maintaining balance of the body and head.
Neurorobotics is the combined study of neuroscience, robotics, and artificial intelligence. It is the science and technology of embodied autonomous neural systems. Neural systems include brain-inspired algorithms, computational models of biological neural networks and actual biological systems. Such neural systems can be embodied in machines with mechanic or any other forms of physical actuation. This includes robots, prosthetic or wearable systems but also, at smaller scale, micro-machines and, at the larger scales, furniture and infrastructures.
In physiology, an efference copy or efferent copy is an internal copy of an outflowing (efferent), movement-producing signal generated by an organism's motor system. It can be collated with the (reafferent) sensory input that results from the agent's movement, enabling a comparison of actual movement with desired movement, and a shielding of perception from particular self-induced effects on the sensory input to achieve perceptual stability. Together with internal models, efference copies can serve to enable the brain to predict the effects of an action.

Spinal locomotion results from intricate dynamic interactions between a central program in lower thoracolumbar spine and proprioceptive feedback from body in the absence of central control by brain as in complete spinal cord injury (SCI). Following SCI, the spinal circuitry below the lesion site does not become silent; rather, it continues to maintain active and functional neuronal properties, although in a modified manner.
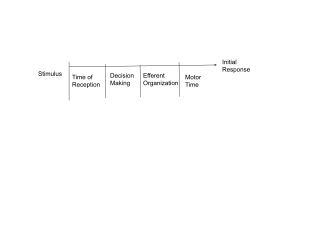
A motor program is an abstract metaphor of the central organization of movement and control of the many degrees of freedom involved in performing an action.p. 182 Signals transmitted through efferent and afferent pathways allow the central nervous system to anticipate, plan or guide movement. Evidence for the concept of motor programs include the following:p. 182

Neuromechanics is an interdisciplinary field that combines biomechanics and neuroscience to understand how the nervous system interacts with the skeletal and muscular systems to enable animals to move. In a motor task, like reaching for an object, neural commands are sent to motor neurons to activate a set of muscles, called muscle synergies. Given which muscles are activated and how they are connected to the skeleton, there will be a corresponding and specific movement of the body. In addition to participating in reflexes, neuromechanical process may also be shaped through motor adaptation and learning.

A spinal interneuron, found in the spinal cord, relays signals between (afferent) sensory neurons, and (efferent) motor neurons. Different classes of spinal interneurons are involved in the process of sensory-motor integration. Most interneurons are found in the grey column, a region of grey matter in the spinal cord.
Postural control refers to the maintenance of body posture in space. The central nervous system interprets sensory input to produce motor output that maintains upright posture. Sensory information used for postural control largely comes from visual, proprioceptive, and vestibular systems. While the ability to regulate posture in vertebrates was previously thought to be a mostly automatic task, controlled by circuits in the spinal cord and brainstem, it is now clear that cortical areas are also involved, updating motor commands based on the state of the body and environment.
Proprioception refers to the sensory information relayed from muscles, tendons, and skin that allows for the perception of the body in space. This feedback allows for more fine control of movement. In the brain, proprioceptive integration occurs in the somatosensory cortex, and motor commands are generated in the motor cortex. In the spinal cord, sensory and motor signals are integrated and modulated by motor neuron pools called central pattern generators (CPGs). At the base level, sensory input is relayed by muscle spindles in the muscle and Golgi tendon organs (GTOs) in tendons, alongside cutaneous sensors in the skin.

Interlimb coordination is the coordination of the left and right limbs. It could be classified into two types of action: bimanual coordination and hands or feet coordination. Such coordination involves various parts of the nervous system and requires a sensory feedback mechanism for the neural control of the limbs. A model can be used to visualize the basic features, the control centre of locomotor movements, and the neural control of interlimb coordination. This coordination mechanism can be altered and adapted for better performance during locomotion in adults and for the development of motor skills in infants. The adaptive feature of interlimb coordination can also be applied to the treatment for CNS damage from stroke and the Parkinson's disease in the future.
