
The mesoderm is the middle layer of the three germ layers that develops during gastrulation in the very early development of the embryo of most animals. The outer layer is the ectoderm, and the inner layer is the endoderm.

The peritoneum is the serous membrane forming the lining of the abdominal cavity or coelom in amniotes and some invertebrates, such as annelids. It covers most of the intra-abdominal organs, and is composed of a layer of mesothelium supported by a thin layer of connective tissue. This peritoneal lining of the cavity supports many of the abdominal organs and serves as a conduit for their blood vessels, lymphatic vessels, and nerves.

A body cavity is any space or compartment, or potential space, in an animal body. Cavities accommodate organs and other structures; cavities as potential spaces contain fluid.
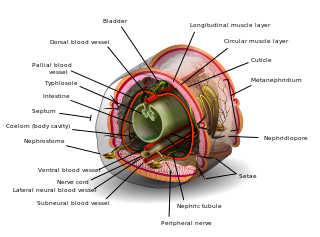
The coelom is the main body cavity in many animals and is positioned inside the body to surround and contain the digestive tract and other organs. In some animals, it is lined with mesothelium. In other animals, such as molluscs, it remains undifferentiated. In the past, and for practical purposes, coelom characteristics have been used to classify bilaterian animal phyla into informal groups.

The mesentery is an organ that attaches the intestines to the posterior abdominal wall and is formed by the double fold of peritoneum. It helps in storing fat and allowing blood vessels, lymphatics, and nerves to supply the intestines, among other functions.

Omphalocele or omphalocoele also called exomphalos, is a rare abdominal wall defect. Beginning at the 6th week of development, rapid elongation of the gut and increased liver size reduces intra abdominal space, which pushes intestinal loops out of the abdominal cavity. Around 10th week, the intestine returns to the abdominal cavity and the process is completed by the 12th week. Persistence of intestine or the presence of other abdominal viscera in the umbilical cord results in an omphalocele.
A germ layer is a primary layer of cells that forms during embryonic development. The three germ layers in vertebrates are particularly pronounced; however, all eumetazoans produce two or three primary germ layers. Some animals, like cnidarians, produce two germ layers making them diploblastic. Other animals such as bilaterians produce a third layer between these two layers, making them triploblastic. Germ layers eventually give rise to all of an animal's tissues and organs through the process of organogenesis.
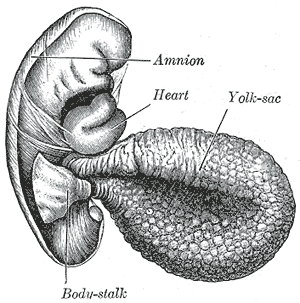
The yolk sac is a membranous sac attached to an embryo, formed by cells of the hypoblast layer of the bilaminar embryonic disc. This is alternatively called the umbilical vesicle by the Terminologia Embryologica (TE), though yolk sac is far more widely used. In humans, the yolk sac is important in early embryonic blood supply, and much of it is incorporated into the primordial gut during the fourth week of embryonic development.
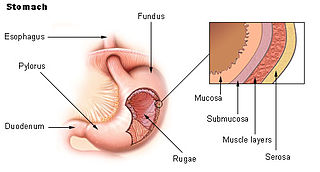
The serous membrane is a smooth tissue membrane of mesothelium lining the contents and inner walls of body cavities, which secrete serous fluid to allow lubricated sliding movements between opposing surfaces. The serous membrane that covers internal organs is called a visceral membrane; while the one that covers the cavity wall is called the parietal membrane. Between the two opposing serosal surfaces is often a potential space, mostly empty except for the small amount of serous fluid.
The primitive node is the organizer for gastrulation in most amniote embryos. In birds it is known as Hensen's node, and in amphibians it is known as the Spemann-Mangold organizer. It is induced by the Nieuwkoop center in amphibians, or by the posterior marginal zone in amniotes including birds.
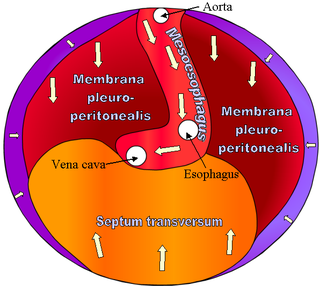
The septum transversum is a thick mass of cranial mesenchyme, formed in the embryo, that gives rise to parts of the thoracic diaphragm and the ventral mesentery of the foregut in the developed human being and other mammals.

The foregut in humans is the anterior part of the alimentary canal, from the distal esophagus to the first half of the duodenum, at the entrance of the bile duct. Beyond the stomach, the foregut is attached to the abdominal walls by mesentery. The foregut arises from the endoderm, developing from the folding primitive gut, and is developmentally distinct from the midgut and hindgut. Although the term “foregut” is typically used in reference to the anterior section of the primitive gut, components of the adult gut can also be described with this designation. Pain in the epigastric region, just below the intersection of the ribs, typically refers to structures in the adult foregut.

The midgut is the portion of the human embryo from which most of the intestines develop. After it bends around the superior mesenteric artery, it is called the "midgut loop". It comprises the portion of the alimentary canal from the end of the foregut at the opening of the bile duct to the hindgut, about two-thirds of the way through the transverse colon.
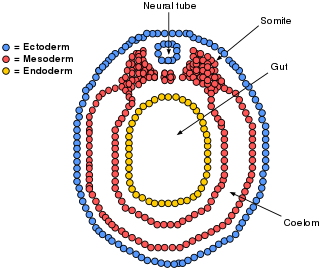
The lateral plate mesoderm is the mesoderm that is found at the periphery of the embryo. It is to the side of the paraxial mesoderm, and further to the axial mesoderm. The lateral plate mesoderm is separated from the paraxial mesoderm by a narrow region of intermediate mesoderm. The mesoderm is the middle layer of the three germ layers, between the outer ectoderm and inner endoderm.
The bilaminar embryonic disc, bilaminar blastoderm or embryonic disc is the distinct two-layered structure of cells formed in an embryo. In the development of the human embryo this takes place by day eight. It is formed when the inner cell mass, also known as the embryoblast, forms a bilaminar disc of two layers, an upper layer called the epiblast and a lower layer called the hypoblast, which will eventually form into fetus. These two layers of cells are stretched between two fluid-filled cavities at either end: the primitive yolk sac and the amniotic sac.

The cloaca is a structure in the development of the urinary and reproductive organs.

Human embryonic development, or human embryogenesis, is the development and formation of the human embryo. It is characterised by the processes of cell division and cellular differentiation of the embryo that occurs during the early stages of development. In biological terms, the development of the human body entails growth from a one-celled zygote to an adult human being. Fertilization occurs when the sperm cell successfully enters and fuses with an egg cell (ovum). The genetic material of the sperm and egg then combine to form the single cell zygote and the germinal stage of development commences. Embryonic development in the human, covers the first eight weeks of development; at the beginning of the ninth week the embryo is termed a fetus. The eight weeks has 23 stages.
The development of the reproductive system is the part of embryonic growth that results in the sex organs and contributes to sexual differentiation. Due to its large overlap with development of the urinary system, the two systems are typically described together as the urogenital or genitourinary system.

In zoology, a mesentery is a membrane inside the body cavity of an animal. The term identifies different structures in different phyla: in vertebrates it is a double fold of the peritoneum enclosing the intestines; in other organisms it forms complete or incomplete partitions of the body cavity, whether that is the coelom or, as in the Anthozoa, the gastrovascular cavity.
