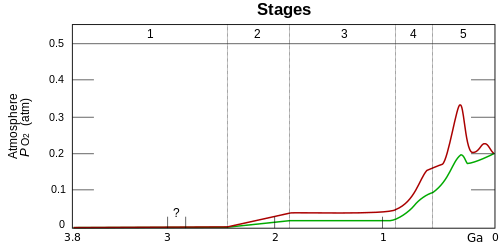
O2 build-up in the Earth's atmosphere. Red and green lines represent the range of the estimates while time is measured in billions of years ago (Ga). Stage 1 (3.85–2.45 Ga): Practically no O2 in the atmosphere. Stage 2 (2.45–1.85 Ga): O2 produced, but absorbed in oceans and seabed rock. Stage 3 (1.85–0.85 Ga): O2 starts to gas out of the oceans, but is absorbed by land surfaces and formation of ozone layer. Stages 4 and 5 (0.85 Ga–present): O2 sinks filled, the gas accumulates.
Oxygen began building up in the prebiotic atmosphere at approximately 2.45 Ga during the Neoarchean-Paleoproterozoic boundary, a paleogeological event known as the Great Oxygenation Event (GOE). The concentrations of O2 attained were less than 10% of today's and probably fluctuated greatly. Around 500Mya a second event known as the Neoproterozoic Oxygenation Event lead to oxygen levels similar or even higher than the present. The increase in oxygen concentrations had wide-ranging and significant impacts on Earth's geochemistry and biosphere. Detailed connections between oxygen and evolution remain elusive.
Importance
Oxygen is both a result of biological activity and a key enabler. Photosynthesis produces oxygen while plants and animals using aerobic respiration consume it. Consequently the evolution of life is closely related to the concentration of available oxygen. Understanding the relationship between oxygen and evolution would aid in seeking evidence of extraterrestrial life in exoplanet data.[4]:252 Oxygen concentration plays a key role in the geochemical composition of sedimentary rocks, making oxygen concentration important for geology and sedimentary rocks important for understanding oxygen concentration over geologic time.[5]
Overview
Due to limitations in measurements, oxygen concentration in the atmosphere or oceans over much of the early parts of Earth's history remains controversial.[6] The consensus view includes these phases:
The timing and character of the oxygenation events have been the subject of many discussions.[6]
Paleo-oxybarometers
Techniques for estimating oxygen at different times in the past are called "paleo-oxybarometers". An ideal technique would rely on trapped gas or fluid in a well-dated rock layer, but such examples are scarce. Most measurements are indirect analysis of oxidation in sedimentary rocks to infer oxygen in ancient atmosphere or oceans. Ocean analysis is especially challenging because tectonicsubduction replaces the ocean floor every 200 million years. Many new techniques have been developed but their analysis and comparison have led to additional debates rather than consensus on the oxygen history of Earth.[8]
Earth's early atmosphere had a very low concentration of oxygen, probably less than 0.001% of present day levels. While details are not well known, measurements of mass-independent fractionation of sulfur in sedimentary sulphides and sulfates rule out significant oxygen before around 2.45Gya.[1] Continuing sources of oxygen during this period would have been photodissociation of water followed by escape the hydrogen product[9] or of sulfur dioxide.[10]
Between 2.45 and 2 Gya, oxygen began to build up in the atmosphere. This time coincided with major changes in geochemistry and biology on the Earth but which changes are causes and which are results are debated.[1]:905 Widespread production of oxygen by cyanobacteria which evolved around this time is suggestive, but substantial evidence suggests that cyanobacteria appeared at least 2.7Ga and perhaps well before that. Geological effects like volcanism, weathering, and burial of chemically altered rocks may be important.[11]:6 Oxygen began to persist in the atmosphere in small quantities about 50 million years before the start of the Great Oxygenation Event.[12] By 800Mya oxygen in the atmosphere was between 1% and 18% of the present atmospheric level.[11]
After the Great Oxygenation Event, the Earth entered a long period of euxinia known as the Boring Billion.[citation needed]} Although the atmosphere had become oxidative with the presence of free oxygen, the oxygen level was still very low (about 0.1%) in both the atmosphere and the ocean. By around 600 Mya during the Neoproterozoic, however, oxygen levels began to rise significantly.[13] During the Cambrian, atmospheric oxygen 5-10% concentration, around half its current value. It rose in pulses above 15% of the atmosphere during the Devonian "Age of the Fishes", reaching 25% (above modern 20%) in the Permo-Carboniferous.[4] This second great oxygenation event has been reported to be associated with the evolution of nitrogen fixation in cyanobacteria,[14] or the rise of more robust eukaryoticphotoautotrophs (i.e. algae) via endosymbiosis and increased phosphorus removal from the ocean.[15]
Atmospheric oxygen is the most conspicuous sign of life on Earth. The evolution of photosynthesis and the rise of oxygen-producing cyanobacteria are major events in evolution. Photosynthetic oxygen eventually accumulated in the atmosphere, transforming both the surface of the planet and the nature of life[11]
Despite these connections, the details are not simple. The evolution of life and the geological history of oxygen share many similar patterns, but the relationship between these two histories remains uncertain. For example, the rise in oxygen concencentration and the rise the maximum size of organism have similar histories, but evidence the oxygen concentration limits size is inconclusive. As more evidence for lower and variable levels of oxygen before the Neoproterozoic has emerged, simple relationships between life and oxygen have been harder to justify.[16]
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.