Histone acetyltransferase p300 also known as p300 HAT or E1A-associated protein p300 also known as EP300 or p300 is an enzyme that, in humans, is encoded by the EP300 gene. It functions as histone acetyltransferase that regulates transcription of genes via chromatin remodeling by allowing histone proteins to wrap DNA less tightly. This enzyme plays an essential role in regulating cell growth and division, prompting cells to mature and assume specialized functions (differentiate), and preventing the growth of cancerous tumors. The p300 protein appears to be critical for normal development before and after birth.
Mothers against decapentaplegic homolog 2 also known as SMAD family member 2 or SMAD2 is a protein that in humans is encoded by the SMAD2 gene. MAD homolog 2 belongs to the SMAD, a family of proteins similar to the gene products of the Drosophila gene 'mothers against decapentaplegic' (Mad) and the C. elegans gene Sma. SMAD proteins are signal transducers and transcriptional modulators that mediate multiple signaling pathways.
Mothers against decapentaplegic homolog 3 also known as SMAD family member 3 or SMAD3 is a protein that in humans is encoded by the SMAD3 gene.

SMAD family member 6, also known as SMAD6, is a protein that in humans is encoded by the SMAD6 gene.

Mothers against decapentaplegic homolog 7 or SMAD7 is a protein that in humans is encoded by the SMAD7 gene.
Smads comprise a family of structurally similar proteins that are the main signal transducers for receptors of the transforming growth factor beta (TGF-B) superfamily, which are critically important for regulating cell development and growth. The abbreviation refers to the homologies to the Caenorhabditis elegans SMA and MAD family of genes in Drosophila.

Activin receptor type-1B is a protein that in humans is encoded by the ACVR1B gene.

Transcription factor jun-B is a protein that in humans is encoded by the JUNB gene. Transcription factor jun-B is a transcription factor involved in regulating gene activity following the primary growth factor response. It binds to the DNA sequence 5'-TGA[CG]TCA-3'.
E3 SUMO-protein ligase PIAS4 is one of several protein inhibitor of activated STAT (PIAS) proteins. It is also known as protein inhibitor of activated STAT protein gamma, and is an enzyme that in humans is encoded by the PIAS4 gene.

C-terminal-binding protein 1 also known as CtBP1 is a protein that in humans is encoded by the CTBP1 gene. CtBP1 is one of two CtBP proteins, the other protein being CtBP2.

Activating transcription factor 2, also known as ATF2, is a protein that, in humans, is encoded by the ATF2 gene.
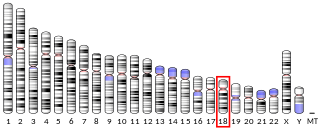
E3 SUMO-protein ligase PIAS2 is an enzyme that in humans is encoded by the PIAS2 gene.

SNW domain-containing protein 1 is a protein that in humans is encoded by the SNW1 gene.

Transcription factor E3 is a protein that in humans is encoded by the TFE3 gene.
Homeobox protein TGIF1 is a protein that, in humans, is encoded by the TGIF1 gene. Alternative splicing has been observed at this locus and eight variants, encoding four distinct isoforms, are described.

MDS1 and EVI1 complex locus protein EVI1 (MECOM) also known as ecotropic virus integration site 1 protein homolog (EVI-1) or positive regulatory domain zinc finger protein 3 (PRDM3) is a protein that in humans is encoded by the MECOM gene. EVI1 was first identified as a common retroviral integration site in AKXD murine myeloid tumors. It has since been identified in a plethora of other organisms, and seems to play a relatively conserved developmental role in embryogenesis. EVI1 is a nuclear transcription factor involved in many signaling pathways for both coexpression and coactivation of cell cycle genes.
Ski-like protein is a protein that in humans is encoded by the SKIL gene.

Serine-threonine kinase receptor-associated protein is an enzyme that in humans is encoded by the STRAP gene.

Krueppel-like factor 10 is a protein that in humans is encoded by the KLF10 gene.
Nuclear factor 1 X-type is a protein that in humans is encoded by the NFIX gene. NFI-X3, a splice variant of NFIX, regulates Glial fibrillary acidic protein and YKL-40 in astrocytes.