Mechanism
Ligand binding
The TGF beta superfamily of ligands includes: bone morphogenetic proteins (BMPs), growth and differentiation factors (GDFs), anti-Müllerian hormone (AMH), Activin, Nodal and TGFβs. [3] Signaling begins with the binding of a TGF beta superfamily ligand to a TGF beta type II receptor. The type II receptor is a serine/threonine receptor kinase, which catalyzes the phosphorylation of the Type I receptor. Each class of ligand binds to a specific type II receptor. [4] In mammals there are seven known type I receptors and five type II receptors. [5]
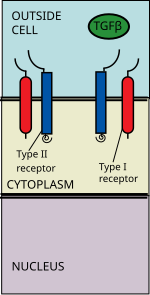
There are three activins: Activin A, Activin B and Activin AB. Activins are involved in embryogenesis and osteogenesis. They also regulate many hormones including pituitary, gonadal and hypothalamic hormones as well as insulin. They are also nerve cell survival factors.
The BMPs bind to the bone morphogenetic protein receptor type-2 (BMPR2). They are involved in a multitude of cellular functions including osteogenesis, cell differentiation, anterior/posterior axis specification, growth, and homeostasis.
The TGFβ family includes: TGFβ1, TGFβ2, TGFβ3. Like the BMPs, TGFβs are involved not only in embryogenesis and cell differentiation, but also in apoptosis and other functions. They bind to TGF-beta receptor type-2 (TGFβR2).
Nodal binds to activin A receptor, type IIB ACVR2B. It can then either form a receptor complex with activin A receptor, type IB (ACVR1B) or with activin A receptor, type IC (ACVR1C). [5]
When the receptor-ligand binding occurs via local action, this is classified as paracrine signalling.
Receptor recruitment and phosphorylation

The TGF beta ligand binds to a type II receptor dimer, which recruits a type I receptor dimer forming a hetero-tetrameric complex with the ligand. [6] These receptors are serine/threonine kinase receptors. They have a cysteine rich extracellular domain, a transmembrane domain, and a cytoplasmic serine/threonine rich domain. The GS domain of the type I receptor consists of a series of about thirty serine-glycine repeats. [7] The binding of a TGFβ family ligand causes the rotation of the receptors so that their cytoplasmic kinase domains are arranged in a catalytically favorable orientation. The Type II receptor phosphorylates serine residues of the Type I receptor, which activates the protein.
SMAD phosphorylation

There are five receptor regulated SMADs: SMAD1, SMAD2, SMAD3, SMAD5, and SMAD9 (sometimes referred to as SMAD8). There are essentially two intracellular pathways involving these R-SMADs. TGFβs, Activins, Nodals and some GDFs are mediated by SMAD2 and SMAD3, while BMPs, AMH and a few GDFs are mediated by SMAD1, SMAD5 and SMAD9. The binding of the R-SMAD to the type I receptor is mediated by a zinc double finger FYVE domain containing protein. Two such proteins that mediate the TGFβ pathway include SARA (the SMAD anchor for receptor activation) and HGS (Hepatocyte growth factor-regulated tyrosine kinase substrate).
SARA is present in an early endosome which, by clathrin-mediated endocytosis, internalizes the receptor complex. [8] SARA recruits an R-SMAD. SARA permits the binding of the R-SMAD to the L45 region of the Type I receptor. [9] SARA orients the R-SMAD such that serine residue on its C-terminus faces the catalytic region of the Type I receptor. The Type I receptor phosphorylates the serine residue of the R-SMAD. Phosphorylation induces a conformational change in the MH2 domain of the R-SMAD and its subsequent dissociation from the receptor complex and SARA. [10]
CoSMAD binding

The now phosphorylated RSMAD has high affinity for coSMAD (e.g. SMAD4) and forms a complex with one. The phosphate group does not act as a docking site for coSMAD, but rather the phosphorylation opens up an amino acid stretch allowing interaction.
Transcription

The phosphorylated RSMAD/coSMAD complex enters the nucleus where it binds transcription promoters/cofactors and causes the transcription of DNA.
Bone morphogenetic proteins cause the transcription of mRNAs involved in osteogenesis, neurogenesis, and ventral mesoderm specification.
TGFβs cause the transcription of mRNAs involved in apoptosis, extracellular matrix neogenesis and immunosuppression. They are also involved in G1 arrest in the cell cycle.
Activin causes the transcription of mRNAs involved in gonadal growth, embryo differentiation and placenta formation.
Nodal causes the transcription of mRNAs involved in left and right axis specification, mesoderm and endoderm induction.