Lipid signaling, broadly defined, refers to any biological cell signaling event involving a lipid messenger that binds a protein target, such as a receptor, kinase or phosphatase, which in turn mediate the effects of these lipids on specific cellular responses. Lipid signaling is thought to be qualitatively different from other classical signaling paradigms (such as monoamineneurotransmission) because lipids can freely diffuse through membranes (see osmosis). One consequence of this is that lipid messengers cannot be stored in vesicles prior to release and so are often biosynthesized "on demand" at their intended site of action. As such, many lipid signaling molecules cannot circulate freely in solution but, rather, exist bound to special carrier proteins in serum.
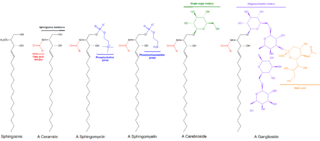
Ceramide contains two hydrophobic ("water-fearing") chains and a neutral headgroup. Consequently, it has limited solubility in water and is restricted within the organelle where it was formed. Also, because of its hydrophobic nature, ceramide readily flip-flops across membranes as supported by studies in membrane models and membranes from red blood cells (erythrocytes).[6] However, ceramide can possibly interact with other lipids to form bigger regions called microdomains which restrict its flip-flopping abilities. This could have immense effects on the signaling functions of ceramide because it is known that ceramide generated by acidic SMase enzymes in the outer leaflet of an organelle membrane may have different roles compared to ceramide that is formed in the inner leaflet by the action of neutral SMase enzymes.[7]
Ceramide mediates many cell-stress responses, including the regulation of programmed cell death (apoptosis) [8] and cell aging (senescence).[9] Numerous research works have focused interest on defining the direct protein targets of action of ceramide. These include enzymes called ceramide-activated Ser-Thr phosphatases (CAPPs), such as proteinphosphatase 1 and 2A (PP1 and PP2A), which were found to interact with ceramide in studies done in a controlled environment outside of a living organism (in vitro).[10] On the other hand, studies in cells have shown that ceramide-inducing agents such as tumor necrosis factor-alpha α (TNFα) and palmitate induce the ceramide-dependent removal of a phosphate group (dephosphorylation) of the retinoblastomagene product RB[11] and the enzymes, protein kinases B (AKT protein family) and C α (PKB and PKCα).[12] Moreover, there is also sufficient evidence which implicates ceramide to the activation of the kinase suppressor of Ras (KSR),[13] PKCζ,[14][15] and cathepsin D.[16]Cathepsin D has been proposed as the main target for ceramide formed in organelles called lysosomes, making lysosomal acidic SMase enzymes one of the key players in the mitochondrial pathway of apoptosis. Ceramide was also shown to activate PKCζ, implicating it to the inhibition of AKT, regulation of the voltage difference between the interior and exterior of the cell (membrane potential) and signaling functions that favor apoptosis.[17]Chemotherapeutic agents such as daunorubicin and etoposide[18][19] enhance the de novo synthesis of ceramide in studies done on mammalian cells. The same results were found for certain inducers of apoptosis particularly stimulators of receptors in a class of lymphocytes (a type of white blood cell) called B-cells.[20] Regulation of the de novo synthesis of ceramide by palmitate may have a key role in diabetes and the metabolic syndrome. Experimental evidence shows that there is substantial increase of ceramide levels upon adding palmitate. Ceramide accumulation activates PP2A and the subsequent dephosphorylation and inactivation of AKT,[21] a crucial mediator in metabolic control and insulinsignaling. This results in a substantial decrease in insulin responsiveness (i.e. to glucose) and in the death of insulin-producing cells in the pancreas called islets of Langerhans.[22] Inhibition of ceramide synthesis in mice via drug treatments or gene-knockout techniques prevented insulin resistance induced by fatty acids, glucocorticoids or obesity.[23]
Ceramide transfer protein (CERT) transports ceramide from ER to the Golgi for the synthesis of SM.[25] CERT is known to bind phosphatidylinositol phosphates, hinting its potential regulation via phosphorylation, a step of the ceramide metabolism that can be enzymatically regulated by protein kinases and phosphatases, and by inositollipid metabolic pathways.[26] Up to date, there are at least 26 distinct enzymes with varied subcellular localizations, that act on ceramide as either a substrate or product. Regulation of ceramide levels can therefore be performed by one of these enzymes in distinct organelles by particular mechanisms at various times.[27]
Sphingosine
Sphingosine (Sph) is formed by the action of ceramidase (CDase) enzymes on ceramide in the lysosome. Sph can also be formed in the extracellular (outer leaflet) side of the plasma membrane by the action of neutral CDase enzyme. Sph then is either recycled back to ceramide or phosphorylated by one of the sphingosine kinase enzymes, SK1 and SK2.[28] The product sphingosine-1-phosphate (S1P) can be dephosphorylated in the ER to regenerate sphingosine by certain S1P phosphatase enzymes within cells, where the salvaged Sph is recycled to ceramide.[29]Sphingosine is a single-chain lipid (usually 18 carbons in length), rendering it to have sufficient solubility in water. This explains its ability to move between membranes and to flip-flop across a membrane. Estimates conducted at physiological pH show that approximately 70% of sphingosine remains in membranes while the remaining 30% is water-soluble.[30] Sph that is formed has sufficient solubility in the liquid found inside cells (cytosol). Thus, Sph may come out of the lysosome and move to the ER without the need for transport via proteins or membrane-enclosed sacs called vesicles. However, its positive charge favors partitioning in lysosomes. It is proposed that the role of SK1 located near or in the lysosome is to ‘trap’ Sph via phosphorylation.[31]
It is important to note that since sphingosine exerts surfactant activity, it is one of the sphingolipids found at lowest cellular levels.[31] The low levels of Sph and their increase in response to stimulation of cells, primarily by activation of ceramidase by growth-inducing proteins such as platelet-derived growth factor and insulin-like growth factor, is consistent with its function as a second messenger. It was found that immediate hydrolysis of only 3 to 10% of newly generated ceramide may double the levels of Sph.[31] Treatment of HL60 cells (a type of leukemia cell line) by a plant-derived organic compound called phorbol ester increased Sph levels threefold, whereby the cells differentiated into white blood cells called macrophages. Treatment of the same cells by exogenous Sph caused apoptosis. A specific protein kinase phosphorylates 14-3-3, otherwise known as sphingosine-dependent protein kinase 1 (SDK1), only in the presence of Sph.[32]
Sph is also known to interact with protein targets such as the protein kinase H homologue (PKH) and the yeast protein kinase (YPK). These targets in turn mediate the effects of Sph and its related sphingoid bases, with known roles in regulating the actincytoskeleton, endocytosis, the cell cycle and apoptosis.[33] It is important to note however that the second messenger function of Sph is not yet established unambiguously.[34]
S1P is probably formed at the inner leaflet of the plasma membrane in response to TNFα and other receptor activity-altering compounds called agonists.[36][37] S1P, being present in low nanomolar concentrations in the cell, has to interact with high-affinity receptors that are capable of sensing their low levels. So far, the only identified receptors for S1P are the high-affinity G protein-coupled receptors (GPCRs), also known as S1P receptors (S1PRs). S1P is required to reach the extracellular side (outer leaflet) of the plasma membrane to interact with S1PRs and launch typical GPCR signaling pathways.[38][39] However, the zwitterionic headgroup of S1P makes it unlikely to flip-flop spontaneously. To overcome this difficulty, the ATP-binding cassette (ABC) transporter C1 (ABCC1) serves as the "exit door" for S1P.[40] On the other hand, the cystic fibrosis transmembrane regulator (CFTR) serves as the means of entry for S1P into the cell.[41] In contrast to its low intracellular concentration, S1P is found in high nanomolar concentrations in serum where it is bound to albumin and lipoproteins.[42] Inside the cell, S1P can induce calcium release independent of the S1PRs—the mechanism of which remains unknown. To date, the intracellular molecular targets for S1P are still unidentified.[31]
The SK1-S1P pathway has been extensively studied in relation to cytokine action, with multiple functions connected to effects of TNFα and IL-1 favoring inflammation. Studies show that knockdown of key enzymes such as S1P lyase and S1P phosphatase increased prostaglandin production, parallel to increase of S1P levels.[37] This strongly suggests that S1P is the mediator of SK1 action and not subsequent compounds. Research done on endothelial and smooth muscle cells is consistent to the hypothesis that S1P has a crucial role in regulating endothelial cell growth, and movement.[43] Recent work on a sphingosine analogue, FTY270, demonstrates its ability to act as a potent compound that alters the activity of S1P receptors (agonist). FTY270 was further verified in clinical tests to have roles in immune modulation, such as that on multiple sclerosis.[44] This highlights the importance of S1P in the regulation of lymphocyte function and immunity. Most of the studies on S1P are used to further understand diseases such as cancer, arthritis and inflammation, diabetes, immune function and neurodegenerative disorders.[31]
Glucosylceramide
Glucosylceramides (GluCer) are the most widely distributed glycosphingolipids in cells serving as precursors for the formation of over 200 known glycosphingolipids. GluCer is formed by the glycosylation of ceramide in an organelle called Golgi via enzymes called glucosylceramide synthase (GCS) or by the breakdown of complex glycosphingolipids (GSLs) through the action of specific hydrolase enzymes. In turn, certain β-glucosidases hydrolyze these lipids to regenerate ceramide.[45][46] GluCer appears to be synthesized in the inner leaflet of the Golgi. Studies show that GluCer has to flip to the inside of the Golgi or transfer to the site of GSL synthesis to initiate the synthesis of complex GSLs. Transferring to the GSL synthesis site is done with the help of a transport protein known as four phosphate adaptor protein 2 (FAPP2) while the flipping to the inside of the Golgi is made possible by the ABC transporter P-glycoprotein, also known as the multi-drug resistance 1 transporter (MDR1).[47] GluCer is implicated in post-Golgi trafficking and drug resistance particularly to chemotherapeutic agents.[48][49] For instance, a study demonstrated a correlation between cellular drug resistance and modifications in GluCer metabolism.[50]
In addition to their role as building blocks of biological membranes, glycosphingolipids have long attracted attention because of their supposed involvement in cell growth, differentiation, and formation of tumors.[31] The production of GluCer from Cer was found to be important in the growth of neurons or brain cells.[51] On the other hand, pharmacological inhibition of GluCer synthase is being considered a technique to avoid insulin resistance.[52]
Ceramide-1-Phosphate
Ceramide-1-phosphate (C1P) is formed by the action of ceramide kinase (CK) enzymes on Cer. C1P carry ionic charge at neutral pH and contain two hydrophobic chains making it relatively insoluble in aqueous environment. Thus, C1P reside in the organelle where it was formed and is unlikely to spontaneously flip-flop across membrane bilayers.[31]
PIP2 binds directly to ion channels and modulates their activity. PIP2 was shown to directly agonizes Inward rectifying potassium channels(Kir).[58] In this regard intact PIP2 signals as a bona fide neurotransmitter-like ligand.[59] PIP2's interaction with many ion channels suggest that the intact form of PIP2 has an important signaling role independent of second messenger signaling.[citation needed]
Second messengers from phosphatidylinositol
Phosphatidylinositol bisphosphate (PIP2) Second Messenger Systems
A general second messenger system mechanism can be broken down into four steps. First, the agonist activates a membrane-bound receptor. Second, the activated G-protein produces a primary effector. Third, the primary effect stimulates the second messenger synthesis. Fourth, the second messenger activates a certain cellular process.
IP3 is soluble and diffuses freely into the cytoplasm. As a second messenger, it is recognized by the inositol triphosphate receptor (IP3R), a Ca2+ channel in the endoplasmic reticulum (ER) membrane, which stores intracellular Ca2+. The binding of IP3 to IP3R releases Ca2+ from the ER into the normally Ca2+-poor cytoplasm, which then triggers various events of Ca2+ signaling. Specifically in blood vessels, the increase in Ca2+ concentration from IP3 releases nitric oxide, which then diffuses into the smooth muscle tissue and causes relaxation.[34]
DAG remains bound to the membrane by its fatty acid "tails" where it recruits and activates both conventional and novel members of the protein kinase C family. Thus, both IP3 and DAG contribute to activation of PKCs.[60][61]
S1P is present at high concentrations in plasma and secreted locally at elevated concentrations at sites of inflammation. It is formed by the regulated phosphorylation of sphingosine. It acts through five dedicated high-affinity G-protein coupled receptors, S1P1 - S1P5. Targeted deletion of S1P1 results in lethality in mice and deletion of S1P2 results in seizures and deafness. Additionally, a mere 3- to 5-fold elevation in serum S1P concentrations induces sudden cardiac death by an S1P3-receptor specific mechanism.
Platelet activating factor (PAF)
PAF is a potent activator of platelet aggregation, inflammation, and anaphylaxis. It is similar to the ubiquitous membrane phospholipidphosphatidylcholine except that it contains an acetyl-group in the SN-2 position and the SN-1 position contains an ether-linkage. PAF signals through a dedicated G-protein coupled receptor, PAFR and is inactivated by PAF acetylhydrolase.
Endocannabinoids
The endogenous cannabinoids, or endocannabinoids, are endogenous lipids that activate cannabinoid receptors. The first such lipid to be isolated was anandamide which is the arachidonoyl amide of ethanolamine. Anandamide is formed via enzymatic release from N-arachidonoyl phosphatidylethanolamine by the N-acyl phosphatidylethanolamine phospholipase D (NAPE-PLD).[62] Anandamide activates both the CB1 receptor, found primarily in the central nervous system, and the CB2 receptor which is found primarily in lymphocytes and the periphery. It is found at very low levels (nM) in most tissues and is inactivated by the fatty acid amide hydrolase. Subsequently, another endocannabinoid was isolated, 2-arachidonoylglycerol, which is produced when phospholipase C releases diacylglycerol which is then converted to 2-AG by diacylglycerol lipase. 2-AG can also activate both cannabinoid receptors and is inactivated by monoacylglycerol lipase. It is present at approximately 100-times the concentration of anandamide in most tissues. Elevations in either of these lipids causes analgesia and anti-inflammation and tissue protection during states of ischemia, but the precise roles played by these various endocannabinoids are still not totally known and intensive research into their function, metabolism, and regulation is ongoing. One saturated lipid from this class, often called an endocannabinoid, but with no relevant affinity for the CB1 and CB 2 receptor is palmitoylethanolamide. This signaling lipid has great affinity for the GRP55 receptor and the PPAR alpha receptor. It has been identified as an anti-inflammatory compound already in 1957, and as an analgesic compound in 1975. Rita Levi-Montalcini first identified one of its biological mechanisms of action, the inhibition of activated mast cells. Palmitoylethanolamide is the only endocannabinoid available on the market for treatment, as a food supplement.
FAHFAs (fatty acid esters of hydroxy fatty acids) are formed in adipose tissue, improve glucose tolerance and also reduce adipose tissue inflammation. Palmitic acid esters of hydroxy-stearic acids (PAHSAs) are among the most bioactive members able to activate G-protein coupled receptors 120.[63] Docosahexaenoic acid ester of hydroxy-linoleic acid (DHAHLA) exert anti-inflammatory and pro-resolving properties.[64]
In biochemistry, a kinase is an enzyme that catalyzes the transfer of phosphate groups from high-energy, phosphate-donating molecules to specific substrates. This process is known as phosphorylation, where the high-energy ATP molecule donates a phosphate group to the substrate molecule. This transesterification produces a phosphorylated substrate and ADP. Conversely, it is referred to as dephosphorylation when the phosphorylated substrate donates a phosphate group and ADP gains a phosphate group. These two processes, phosphorylation and dephosphorylation, occur four times during glycolysis.
Sphingolipids are a class of lipids containing a backbone of sphingoid bases, which are a set of aliphatic amino alcohols that includes sphingosine. They were discovered in brain extracts in the 1870s and were named after the mythological sphinx because of their enigmatic nature. These compounds play important roles in signal transduction and cell recognition. Sphingolipidoses, or disorders of sphingolipid metabolism, have particular impact on neural tissue. A sphingolipid with a terminal hydroxyl group is a ceramide. Other common groups bonded to the terminal oxygen atom include phosphocholine, yielding a sphingomyelin, and various sugar monomers or dimers, yielding cerebrosides and globosides, respectively. Cerebrosides and globosides are collectively known as glycosphingolipids.
Sphingomyelin is a type of sphingolipid found in animal cell membranes, especially in the membranous myelin sheath that surrounds some nerve cell axons. It usually consists of phosphocholine and ceramide, or a phosphoethanolamine head group; therefore, sphingomyelins can also be classified as sphingophospholipids. In humans, SPH represents ~85% of all sphingolipids, and typically make up 10–20 mol % of plasma membrane lipids.
Ceramides are a family of waxy lipid molecules. A ceramide is composed of sphingosine and a fatty acid joined by an amide bond. Ceramides are found in high concentrations within the cell membrane of eukaryotic cells, since they are component lipids that make up sphingomyelin, one of the major lipids in the lipid bilayer. Contrary to previous assumptions that ceramides and other sphingolipids found in cell membrane were purely supporting structural elements, ceramide can participate in a variety of cellular signaling: examples include regulating differentiation, proliferation, and programmed cell death (PCD) of cells.
Sphingosine kinase (SphK) is a conserved lipid kinase that catalyzes formation sphingosine-1-phosphate (S1P) from the precursor sphingolipid sphingosine. Sphingolipid metabolites, such as ceramide, sphingosine and sphingosine-1-phosphate, are lipid second messengers involved in diverse cellular processes. There are two forms of SphK, SphK1 and SphK2. SphK1 is found in the cytosol of eukaryotic cells, and migrates to the plasma membrane upon activation. SphK2 is localized to the nucleus.
Sphingosine-1-phosphate (S1P) is a signaling sphingolipid, also known as lysosphingolipid. It is also referred to as a bioactive lipid mediator. Sphingolipids at large form a class of lipids characterized by a particular aliphatic aminoalcohol, which is sphingosine.
Sphingosine kinase 1 is an enzyme that in humans is encoded by the SPHK1 gene.
Ceramidase is an enzyme which cleaves fatty acids from ceramide, producing sphingosine (SPH) which in turn is phosphorylated by a sphingosine kinase to form sphingosine-1-phosphate (S1P).
Sphingosine-1-phosphate receptor 1, also known as endothelial differentiation gene 1 (EDG1) is a protein that in humans is encoded by the S1PR1 gene. S1PR1 is a G-protein-coupled receptor which binds the bioactive signaling molecule sphingosine 1-phosphate (S1P). S1PR1 belongs to a sphingosine-1-phosphate receptor subfamily comprising five members (S1PR1-5). S1PR1 was originally identified as an abundant transcript in endothelial cells and it has an important role in regulating endothelial cell cytoskeletal structure, migration, capillary-like network formation and vascular maturation. In addition, S1PR1 signaling is important in the regulation of lymphocyte maturation, migration and trafficking.
Sphingosine-1-phosphate receptor 3 also known as S1PR3 is a human gene which encodes a G protein-coupled receptor which binds the lipid signaling molecule sphingosine 1-phosphate (S1P). Hence this receptor is also known as S1P3.
Sphingosine-1-phosphate receptor 2, also known as S1PR2 or S1P2, is a human gene which encodes a G protein-coupled receptor which binds the lipid signaling molecule sphingosine 1-phosphate (S1P).
The enzyme phosphatidate phosphatase (PAP, EC 3.1.3.4) is a key regulatory enzyme in lipid metabolism, catalyzing the conversion of phosphatidate to diacylglycerol:
G-protein coupled receptor 3 is a protein that in humans is encoded by the GPR3 gene. The protein encoded by this gene is a member of the G protein-coupled receptor family of transmembrane receptors and is involved in signal transduction.
In enzymology, a ceramide kinase, also abbreviated as CERK, is an enzyme that catalyzes the chemical reaction:
Lipid phosphate phosphohydrolase 1 also known as phosphatidic acid phosphatase 2a is an enzyme that in humans is encoded by the PPAP2A gene.
Phosphatidylinositol-4-phosphate 5-kinase type-1 alpha is an enzyme that in humans is encoded by the PIP5K1A gene.
Sphingosine kinase 2 is a protein that in humans is encoded by the SPHK2 gene.
Ceramide synthase 2, also known as LAG1 longevity assurance homolog 2 or Tumor metastasis-suppressor gene 1 protein is an enzyme that in humans is encoded by the CERS2 gene.
Ceramide synthase 5 (CerS5) is the enzyme encoded in humans by the CERS5 gene.
Cells destined for apoptosis release molecules referred to as find-me signals. These signal molecules are used to attract phagocytes which engulf and eliminate damaged cells. Find-me signals are typically released by the apoptotic cells while the cell membrane remains intact. This ensures that the phagocytic cells are able to remove the dying cells before their membranes are compromised. A leaky membrane leads to secondary necrosis which may cause additional inflammation, therefore, it is best to remove dying cells before this occurs. One cell is capable of releasing multiple find-me signals. Should a cell lack the ability to release its find-me signal, other cells may release additional find-me signals to overcome the discrepancy.
↑ Marchesini, N.; Hannun, Y. A. (2004). "Acid and neutral sphingomyelinases: roles and mechanisms of regulation". Biochem. Cell Biol. 82 (1): 27–44. doi:10.1139/o03-091. PMID15052326.
↑ Obeid, L. M., Linardic, C. M., Karolak, L. A. & Hannun, Y. A. (1993) Programmed cell death induced by ceramide. Science. 259, 1769–1771 .
↑ Hamaguchi, A.; etal. (2003). "A sphingosine-dependent protein kinase that specifically phosphorylates 14-3-3 (SDK1) is identified as the kinase domain of PKC: a preliminary note. Biochemical and". Biophys. Res. Comm. 307 (3): 589–594. doi:10.1016/S0006-291X(03)01070-2. PMID12893264.
↑ Smith, E. R.; Merrill, A. H.; Obeid, L. M.; Hannun, Y. A. (2000). "Effects of Sphingosine and Other Sphingolipids on Protein Kinase C". Sphingolipid Metabolism and Cell Signaling, Part B. Methods in Enzymology. Vol.312. pp.361–373. doi:10.1016/S0076-6879(00)12921-0. ISBN9780121822132. PMID11070884.
↑ Bandhuvula, P.; Saba, J. D. (2007). "Sphingosine-1-phosphate lyase in immunity and cancer: silencing the siren". Trends Mol. Med. 13 (5): 210–217. doi:10.1016/j.molmed.2007.03.005. PMID17416206.
↑ Taha, T. A.; Argraves, K. M.; Obeid, L. M. (2004). "Sphingosine-1-phosphate receptors: receptor specificity versus functional redundancy". Biochim. Biophys. Acta. 1682 (1–3): 48–55. doi:10.1016/j.bbalip.2004.01.006. PMID15158755.
↑ Okajima, F. (2002). "Plasma lipoproteins behave as carriers of extracellular sphingosine 1-phosphate: is this an atherogenic mediator or an anti-atherogenic mediator?". Biochim. Biophys. Acta. 1582 (1–3): 132–137. doi:10.1016/s1388-1981(02)00147-6. PMID12069820.
↑ Peters, S. L.; Alewijnse, A. E. (2007). "Sphingosine-1-phosphate signaling in the cardiovascular system". Current Opinion in Pharmacology. 7 (2): 186–192. doi:10.1016/j.coph.2006.09.008. PMID17280869.
↑ Radin, N. S., Shayman, J.A. & Inokuchi, J.-I. Metabolic effects of inhibiting glucosylceramide synthesis with PDMP and other substances. Adv. Lipid Res.26, 183–211
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.