Some microorganisms use retinal to convert light into metabolic energy. One study suggests that approximately three billion years ago, most living organisms on Earth used retinal, rather than chlorophyll, to convert sunlight into energy. Because retinal absorbs mostly green light and transmits purple light, this gave rise to the Purple Earth hypothesis.[2]
Retinal itself is considered to be a form of vitamin A when eaten by an animal. There are many forms of vitamin A, all of which are converted to retinal, which cannot be made without them. The number of different molecules that can be converted to retinal varies from species to species. Retinal was originally called retinene,[3] and was renamed[4] after it was discovered to be vitamin A aldehyde.[5][6]
Vertebrate animals ingest retinal directly from meat, or they produce retinal from carotenoids – either from α-carotene or β-carotene – both of which are carotenes. They also produce it from β-cryptoxanthin, a type of xanthophyll. These carotenoids must be obtained from plants or other photosynthetic organisms. No other carotenoids can be converted by animals to retinal. Some carnivores cannot convert any carotenoids at all. The other main forms of vitamin A – retinol and a partially active form, retinoic acid – may both be produced from retinal.
Invertebrates such as insects and squid use hydroxylated forms of retinal in their visual systems, which derive from conversion from other xanthophylls.
Vitamin A metabolism
Living organisms produce retinal by irreversible oxidative cleavage of carotenoids.[7]
Just as carotenoids are the precursors of retinal, retinal is the precursor of the other forms of vitamin A. Retinal is interconvertible with retinol, the transport and storage form of vitamin A:
Retinoic acid, sometimes called vitamin A acid, is an important signaling molecule and hormone in vertebrate animals.
Vision
Retinal is a conjugated chromophore. In the Vertebrate eyes, retinal begins in an 11-cis-retinal configuration, which — upon capturing a photon of the correct wavelength — straightens out into an all-trans-retinal configuration. This configuration change pushes against an opsin protein in the retina, which triggers a chemical signaling cascade, which results in perception of light or images by the brain. The absorbance spectrum of the chromophore depends on its interactions with the opsin protein to which it is bound, so that different retinal-opsin complexes will absorb photons of different wavelengths (i.e., different colors of light).
Opsins
An opsin protein surrounds a molecule of 11-cis retinal, awaiting the arrival of a photon. Once the retinal molecule captures a photon, its configuration change causes it to push against the surrounding opsin protein which may cause the opsin to send a chemical signal to the brain indicating that light has been detected. Retinal is then converted back to its 11-cis configuration by ATP phosphorylation, and the cycle begins again.Animal GPCR rhodopsin (rainbow-colored) embedded in a lipid bilayer (heads red and tails blue) with transducin below it. Gtα is colored red, Gtβ blue, and Gtγ yellow. There is a bound GDP molecule in the Gtα-subunit and a bound retinal (black) in the rhodopsin. The N-terminus terminus of rhodopsin is red and the C-terminus blue. Anchoring of transducin to the membrane has been drawn in black.
Retinal is bound to opsins, which are G protein-coupled receptors (GPCRs).[14][15] Opsins, like other GPCRs, have seven transmembrane alpha-helices connected by six loops. They are found in the photoreceptor cells in the retina of eye. The opsin in the vertebrate rod cells is rhodopsin. The rods form disks, which contain the rhodopsin molecules in their membranes and which are entirely inside of the cell. The N-terminus head of the molecule extends into the interior of the disk, and the C-terminus tail extends into the cytoplasm of the cell. The opsins in the cone cells are OPN1SW, OPN1MW, and OPN1LW. The cones form incomplete disks that are part of the plasma membrane, so that the N-terminus head extends outside of the cell. In opsins, retinal binds covalently to a lysine[16] in the seventh transmembrane helix[17][18][19] through a Schiff base.[20][21] Forming the Schiff base linkage involves removing the oxygen atom from retinal and two hydrogen atoms from the free amino group of lysine, giving H2O. Retinylidene is the divalent group formed by removing the oxygen atom from retinal, and so opsins have been called retinylidene proteins.
Although mammals use retinal exclusively as the opsin chromophore, other groups of animals additionally use four chromophores closely related to retinal: 3,4-didehydroretinal (vitamin A2), (3R)-3-hydroxyretinal, (3S)-3-hydroxyretinal (both vitamin A3), and (4R)-4-hydroxyretinal (vitamin A4). Many fish and amphibians use 3,4-didehydroretinal, also called dehydroretinal. With the exception of the dipteran suborder Cyclorrhapha (the so-called higher flies), all insects examined use the (R)-enantiomer of 3-hydroxyretinal. The (R)-enantiomer is to be expected if 3-hydroxyretinal is produced directly from xanthophyll carotenoids. Cyclorrhaphans, including Drosophila, use (3S)-3-hydroxyretinal.[28][29]Firefly squid have been found to use (4R)-4-hydroxyretinal.
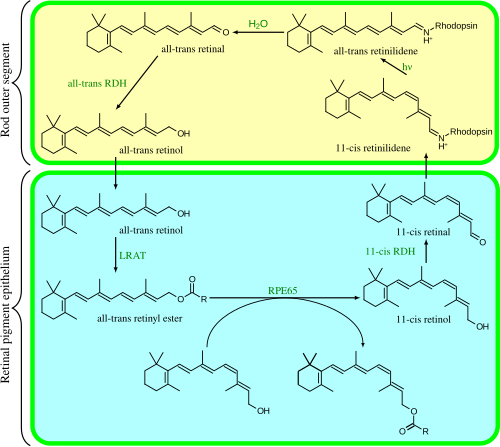
The visual cycle is a circular enzymatic pathway, which is the front-end of phototransduction. It regenerates 11-cis-retinal. For example, the visual cycle of mammalian rod cells is as follows:
RPE65 isomerohydrolases are homologous with beta-carotene monooxygenases;[7] the homologous ninaB enzyme in Drosophila has both retinal-forming carotenoid-oxygenase activity and all-trans to 11-cis isomerase activity.[32]
All-trans-retinal is also an essential component of microbial opsins such as bacteriorhodopsin, channelrhodopsin, and halorhodopsin, which are important in bacterial and archaealanoxygenic photosynthesis. In these molecules, light causes the all-trans-retinal to become 13-cis retinal, which then cycles back to all-trans-retinal in the dark state. These proteins are not evolutionarily related to animal opsins and are not GPCRs; the fact that they both use retinal is a result of convergent evolution.[33]
↑Attwood, T. K.; Findlay, J. B. C. (1994). "Fingerprinting G-protein-coupled receptors". Protein Engineering, Design and Selection. 7 (2): 195–203. doi:10.1093/protein/7.2.195. PMID8170923.
↑Hargrave, P. A.; McDowell, J. H.; Curtis, Donna R.; Wang, Janet K.; Juszczak, Elizabeth; Fong, Shao-Ling; Mohana Rao, J. K.; Argos, P. (1983). "The structure of bovine rhodopsin". Biophysics of Structure and Mechanism. 9 (4): 235–244. doi:10.1007/BF00535659. PMID6342691. S2CID20407577.
↑Kumbalasiri T, Rollag MD, Isoldi MC, Castrucci AM, Provencio I (March 2007). "Melanopsin triggers the release of internal calcium stores in response to light". Photochemistry and Photobiology. 83 (2): 273–279. doi:10.1562/2006-07-11-RA-964. PMID16961436. S2CID23060331.
↑Seki, Takaharu; Isono, Kunio; Ito, Masayoshi; Katsuta, Yuko (1994). "Flies in the Group Cyclorrhapha Use (3S)-3-Hydroxyretinal as a Unique Visual Pigment Chromophore". European Journal of Biochemistry. 226 (2): 691–696. doi:10.1111/j.1432-1033.1994.tb20097.x. PMID8001586.
Barlow, H.B.; Levick, W.R.; Yoon, M. (1971). "Responses to single quanta of light in retinal ganglion cells of the cat". Vision Research. 11 (Supplement 3): 87–101. doi:10.1016/0042-6989(71)90033-2. PMID5293890.
Sadekar, Sumedha; Raymond, Jason; Blankenship, Robert E. (2006). "Conservation of Distantly Related Membrane Proteins: Photosynthetic Reaction Centers Share a Common Structural Core". Molecular Biology and Evolution. 23 (11): 2001–2007. doi:10.1093/molbev/msl079. PMID16887904.
Send, Robert; Sundholm, Dage (2007). "Stairway to the conical intersection: A computational study of retinal isomerization". Journal of Physical Chemistry A. 111 (36): 8766–8773. Bibcode:2007JPCA..111.8766S. doi:10.1021/jp073908l. PMID17713894.
Su, Chih-Ying; Luo, Dong-Gen; Terakita, Akihisa; Shichida, Yoshinori; Liao, Hsi-Wen; Kazmi, Manija A.; Sakmar, Thomas P.; Yau, King-Wai (2006). "Parietal-Eye Phototransduction Components and Their Potential Evolutionary Implications". Science. 311 (5767): 1617–1621. Bibcode:2006Sci...311.1617S. doi:10.1126/science.1123802. PMID16543463. S2CID28604455.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.