Bacteriorhodopsin is a light-driven H+ ion transporter found in some Haloarchaea, most notably Halobacterium salinarum (formerly known as syn. H.halobium). The proton-motive force generated by the protein is used by ATP synthase to generate adenosine triphosphate (ATP). By expressing Bacteriorhodopsin, the archaea cells are able to synthesise ATP in the absence of a carbon source.[4][5]
Structure
A bacteriorhodopsin trimer, showing the approximate positions of the extracellular and cytoplasmic sides of the membrane (red and blue lines respectively)
Bacteriorhodopsin is a 27kDaintegral membrane protein usually found in two-dimensional crystalline patches known as "purple membrane", which can occupy almost 50% of the surface area of the archaeal cell. The repeating element of the hexagonal lattice is composed of three identical protein chains, each rotated by 120 degrees relative to the others.[6] Each monomer has seven transmembrane alpha helices and an extracellular-facing, two-stranded beta sheet.[7][8]
Bacteriorhodopsin molecule is purple and is most efficient at absorbing green light (in the wavelength range 500-650 nm). In the native membrane, the protein has a maximum absorbance at 553 nm, however addition of detergent disrupts the trimeric form, leading a loss of exciton coupling between the chromophores, and the monomeric form consequently has an absorption maximum of 568 nm.[13][14]
Bacteriorhodopsin has a broad excitation spectrum. For a detection wavelength between 700 and 800nm, it has an appreciable detected emission for excitation wavelengths between 470nm and 650nm (with a peak at 570nm).[15] When pumped at 633nm, the emission spectrum has appreciable intensity between 650nm and 850nm.[16]
Mechanism
Photocycle overview
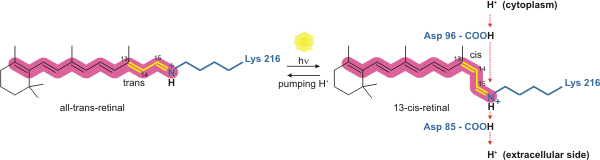
Bacteriorhodopsin is a light-driven proton pump. It is the retinal molecule that changes its isomerization state from all-trans to 13-cis when it absorbs a photon. The surrounding protein responds to the change in the chromophore shape, by undergoing an ordered sequence of conformational changes (collectively known as the photocycle).[17] The conformational changes alter the pKa values of conserved amino acids in the core of the protein, including Asp85, Asp96 and the Schiff base N atom (Lys216). These sequential changes in acid dissociation constant, result in the transfer of one proton from the intracellular side to the extracellular side of the membrane for each photon absorbed by the chromophore.
The bacteriorhodopsin photocycle consists of nine distinct stages, starting from the ground or resting state, which is denoted 'bR'. The intermediates are identified by single letters and may be distinguished by their absorption spectra.[18] The nine stages are:
bR + photon → K ⇌ L ⇌ M1⇌ M2⇌ M2' ⇌ N ⇌ N' ⇌ O ⇌ bR[18]
Ground state + photon → K state → L state
Conformational change, paired stereogram. The orange molecule is all-trans retinal and the red molecule is 13-cis retinal.
Bacteriorhodopsin in the ground state absorbs a photon and the retinal changes isomerization from all-trans 15-anti to the strained 13-cis 15-anti in the K state. The isomerisation reaction is fast and occurs in less than 1ps. The retinal adopts a less strained conformation to form the L intermediate.
L state → M1 state
Asp85 accepts a proton from the Schiff base N atom. In the M1 intermediate, neither the Schiff base nor Asp85 are charged.
M1 state → M2 state
The Schiff base rotates away from the extracellular side of the protein towards the cytoplasmic side, in preparation to accept a new proton.
M2 state → M2' state
A proton is released from Glu204 and Glu194 to the extracellular medium.
M2' state → N state
The retinal Schiff base accepts a proton from Asp96. In the N state, both Asp96 and the Schiff base are charged.
N state → N' state
Asp96 accepts a proton from the cytoplasmic side of the membrane and becomes uncharged.
N' state → O state
Retinal reisomerizes to the all-trans state.
O state → ground state
Asp85 transfers a proton to Glu194 and Glu204[19][20] on the extracellular face of the protein.
Bacteriorhodopsin is similar to vertebraterhodopsins, the pigments that sense light in the retina. Rhodopsins also contain retinal; however, the functions of rhodopsin and bacteriorhodopsin are different, and there is limited similarity in their amino acid sequences. Both rhodopsin and bacteriorhodopsin belong to the 7TM receptor family of proteins, but rhodopsin is a G protein-coupled receptor and bacteriorhodopsin is not. In the first use of electron crystallography to obtain an atomic-level protein structure, the structure of bacteriorhodopsin was resolved in 1990.[22] It was then used as a template to build models of G protein-coupled receptors before crystallographic structures were also available for these proteins. It has been excessively studied on both mica[23][24] and glass substrates using Atomic force microscopy and Femtosecond crystallography.[25]
All other phototrophic systems in bacteria, algae, and plants use chlorophylls or bacteriochlorophylls rather than bacteriorhodopsin. These also produce a proton gradient, but in a quite different and more indirect way involving an electron transfer chain consisting of several other proteins. Furthermore, chlorophylls are aided in capturing light energy by other pigments known as "antennas"; these are not present in bacteriorhodopsin-based systems. It is possible that phototrophy independently evolved at least twice, once in bacteria and once in archaea.
↑ Pebay-Peroua E, Rummel G, Rosenbusch JP, Landau EM (1997). "X-ray structure of bacteriorhodopsin at 2.5 Å from microcrystals grown in lipidic cubic phases". Science. 277 (5332): 1676–1681. doi:10.1126/science.277.5332.1676. PMID9287223.
↑ Luecke H, Schobert B, Richter HT, Cartailler JP, Lanyi JK (1999). "Structure of bacteriorhodopsin at 1.55 Å resolution". Journal of Molecular Biology. 291 (4): 899–911. doi:10.1006/jmbi.1999.3027. PMID10452895.
↑ Oesterhelt, Dieter (1982). "[3] Reconstitution of the retinal proteins bacteriorhodopsin and halorhodopsin". Reconstitution of the retinal proteins bacteriorhodopsin and halorhodopsin. Methods in Enzymology. Vol.88. pp.10–17. doi:10.1016/0076-6879(82)88006-3. ISBN9780121819880.
↑ Pescitelli G, Woody RW (2012). "The Exciton Origin of the Visible Circular Dichroism Spectrum of Bacteriorhodopsin". Journal of Physical Chemistry B. 116 (23): 6751–6763. doi:10.1021/jp212166k. PMID22329810.
↑ Schenkl, Selma; Zgrablic, Goran; Portuondo-Campa, Erwin; Haacke, Stefan; Chergui, Majed (2007). "On the excitation wavelength dependence of the fluorescence of bacteriorhodopsin". Chemical Physics Letters. 441 (4–6): 322–326. Bibcode:2007CPL...441..322S. doi:10.1016/j.cplett.2007.04.086.
↑ Ohtani, H.; Tsukamoto, Y.; Sakoda, Y.; Hamaguchi, H. (1995). "Fluorescence spectra of bacteriorhodopsin and the intermediates O and Q at room temperature". FEBS Lett. 359 (1): 65–68. doi:10.1016/0014-5793(94)01440-c. PMID7851532.
↑ Dioumaev, A. K.; Richter, H. T.; Brown, L. S.; Tanio, M.; Tuzi, S.; Saito, H.; Kimura, Y.; Needleman, R.; Lanyi, J. K. (1998). "Existence of a proton transfer chain in bacteriorhodopsin: Participation of Glu-194 in the release of protons to the extracellular surface". Biochemistry. 37 (8): 2496–2906. doi:10.1021/bi971842m. PMID9485398.
↑ Balashov, S. P.; Lu, M.; Imasheva, E. S.; Govindjee, R.; Ebrey, T. G.; Othersen b, 3rd; Chen, Y.; Crouch, R. K.; Menick, D. R. (1999). "The proton release group of bacteriorhodopsin controls the rate of the final step of its photocycle at low pH". Biochemistry. 38 (7): 2026–2039. doi:10.1021/bi981926a. PMID10026285.{{cite journal}}: CS1 maint: numeric names: authors list (link)
↑ Henderson R, Baldwin JM, Ceska TA, Zemlin F, Beckmann E, Downing KH (1990). "Model for the structure of bacteriorhodopsin based on high-resolution electron cryo-microscopy". J Mol Biol. 213 (4): 899–929. doi:10.1016/S0022-2836(05)80271-2. PMID2359127.{{cite journal}}: CS1 maint: multiple names: authors list (link)
1 2 Nishikawa, T.; Murakami, M. (2005). "Crystal structure of the 13-cis isomer of bacteriorhodopsin in the dark-adapted state". J. Mol. Biol. 352 (2): 319–328. doi:10.1016/j.jmb.2005.07.021. PMID16084526. PDB ID: 1X0S.
1 2 Image created with RasTop (Molecular Visualization Software).
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.