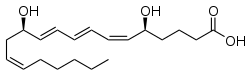
LTA4 Note the four double bonds, three of them conjugated. This is a common property of A4, B4, C4, D4, and E4.LTB4LTC4 is a cysteinyl leukotriene, as are D4 and E4.LTD4LTE4
Leukotrienes use lipid signaling to convey information to either the cell producing them (autocrine signaling) or neighboring cells (paracrine signaling) in order to regulate immune responses. The production of leukotrienes is usually accompanied by the production of histamine and prostaglandins, which also act as inflammatory mediators.[4]
One of their roles (specifically, leukotriene D4) is to trigger contractions in the smooth muscles lining the bronchioles; their overproduction is a major cause of inflammation in asthma and allergic rhinitis.[5]Leukotriene antagonists are used to treat these disorders by inhibiting the production or activity of leukotrienes.[6]
History and name
The name leukotriene, introduced by Swedish biochemist Bengt Samuelsson in 1979, comes from the words leukocyte and triene (indicating the compound's three conjugated double bonds). What would be later named leukotriene C, "slow reaction smooth muscle-stimulating substance" (SRS) was originally described between 1938 and 1940 by Feldberg and Kellaway.[7][8][9] The researchers isolated SRS from lung tissue after a prolonged period following exposure to snake venom and histamine.[9]
Types
Cysteinyl leukotrienes
LTC4, LTD4, LTE4 and LTF4 are often called cysteinyl leukotrienes due to the presence of the amino acid cysteine in their structure. The cysteinyl leukotrienes make up the slow-reacting substance of anaphylaxis (SRS-A). LTF4, like LTD4, is a metabolite of LTC4, but, unlike LTD4, which lacks the glutamic residue of glutathione, LTF4 lacks the glycine residue of glutathione.[10]
LTB4 is synthesized in vivo from LTA4 by the enzyme LTA4 hydrolase. Its primary function is to recruit neutrophils to areas of tissue damage, though it also helps promote the production of inflammatory cytokines by various immune cells. Drugs that block the actions of LTB4 have shown some efficacy in slowing the progression of neutrophil-mediated diseases.[11]
LTG4
There has also been postulated the existence of LTG4, a metabolite of LTE4 in which the cysteinyl moiety has been oxidized to an alpha-keto-acid (i.e.—the cysteine has been replaced by a pyruvate). Very little is known about this putative leukotriene.[citation needed]
LTB5
Leukotrienes originating from the omega-3 class eicosapentanoic acid (EPA) have diminished inflammatory effects. In human subjects whose diets have been supplemented with eicosapentaenoic acid, leukotrine B5, along with leukotrine B4, is produced by neutrophils.[12] LTB5 induces aggregation of rat neutrophils, chemokinesis of human polymorphonuclear neutrophils (PMN), lysosomal enzyme release from human PMN and potentiation of bradykinin-induced plasma exudation, although compared to LTB4, it has at least 30 times less potency.[13]
Biochemistry
Synthesis
Eicosanoid synthesis. (Leukotrienes at right.)
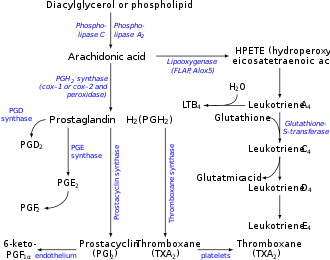
Leukotrienes are synthesized in the cell from arachidonic acid by arachidonate 5-lipoxygenase. The catalytic mechanism involves the insertion of an oxygen moiety at a specific position in the arachidonic acid backbone.[citation needed]
5-Lipoxygenase (5-LO) uses FLAP to convert arachidonic acid into 5-hydroperoxyeicosatetraenoic acid (5-HPETE), which spontaneously reduces to 5-hydroxyeicosatetraenoic acid (5-HETE). The enzyme 5-LO acts again on 5-HETE to convert it into leukotriene A4 (LTA4), an unstable epoxide. 5-HETE can be further metabolized to 5-oxo-ETE and 5-oxo-15-hydroxy-ETE, all of which have pro-inflammatory actions similar but not identical to those of LTB4 and mediated not by LTB4 receptors but rather by the OXE receptor (see 5-Hydroxyeicosatetraenoic acid and 5-Oxo-eicosatetraenoic acid).[14][15]
In cells equipped with LTA hydrolase, such as neutrophils and monocytes, LTA4 is converted to the dihydroxy acid leukotriene LTB4, which is a powerful chemoattractant for neutrophils acting at BLT1 and BLT2 receptors on the plasma membrane of these cells.[citation needed]
In cells that express LTC4 synthase, such as mast cells and eosinophils, LTA4 is conjugated with the tripeptide glutathione to form the first of the cysteinyl-leukotrienes, LTC4. Outside the cell, LTC4 can be converted by ubiquitous enzymes to form successively LTD4 and LTE4, which retain biological activity.[citation needed]
The cysteinyl-leukotrienes act at their cell-surface receptors CysLT1 and CysLT2 on target cells to contract bronchial and vascular smooth muscle, to increase permeability of small blood vessels, to enhance secretion of mucus in the airway and gut, and to recruit leukocytes to sites of inflammation.[citation needed]
Both LTB4 and the cysteinyl-leukotrienes (LTC4, LTD4, LTE4) are partly degraded in local tissues, and ultimately become inactive metabolites in the liver.[citation needed]
Leukotrienes are very important agents in the inflammatory response. Some such as LTB4 have a chemotactic effect on migrating neutrophils, and as such help to bring the necessary cells to the tissue. Leukotrienes also have a powerful effect in bronchoconstriction and increase vascular permeability.[17]
Infiltration of inflammatory cells in the airway wall
Role of cysteinyl leukotrienes
Cysteinyl leukotriene receptors CYSLTR1 and CYSLTR2 are present on mast cells, eosinophil, and endothelial cells. During cysteinyl leukotriene interaction, they can stimulate proinflammatory activities such as endothelial cell adherence and chemokine production by mast cells. As well as mediating inflammation, they induce asthma and other inflammatory disorders, thereby reducing the airflow to the alveoli. The levels of cysteinyl leukotrienes, along with 8-isoprostane, have been reported to be increased in the EBC of patients with asthma, correlating with disease severity.[19] Cysteinyl leukotrienes may also play a role in adverse drug reactions in general and in contrast medium induced adverse reactions in particular.[20]
↑ Loick, H.; Theissen, J. (1994). "Die Eicosanoide als Mediatoren beim ARDS" [Eicosanoids as mediators in ARDS]. Anästhesiologie, Intensivmedizin, Notfallmedizin, Schmerztherapie (in German). 29 (1): 3–9. doi:10.1055/s-2007-996677. PMID8142566.
↑ Scott JP, Peters-Golden M (September 2018). "Antileukotriene agents for the treatment of lung disease". Am. J. Respir. Crit. Care Med. 188 (5): 538–544. doi:10.1164/rccm.201301-0023PP. PMID23822826.
↑ Manev, Radmila; Manev, Hari (2004). "5-Lipoxygenase as a Putative Link Between Cardiovascular and Psychiatric Disorders". Critical Reviews in Neurobiology. 16 (1–2): 181–6. doi:10.1615/CritRevNeurobiol.v16.i12.190. PMID15581413.
↑ Böhm, Ingrid; Speck, Ulrich; Schild, Hans (2005). "A possible role for cysteinyl-leukotrienes in non-ionic contrast media induced adverse reactions". European Journal of Radiology. 55 (3): 431–6. doi:10.1016/j.ejrad.2005.01.007. PMID16129253.
Bailey, J. Martyn (1985) Prostaglandins, leukotrienes, and lipoxins: biochemistry, mechanism of action, and clinical applications Plenum Press, New York, ISBN0-306-41980-7
Lipkowitz, Myron A. and Navarra, Tova (2001) The Encyclopedia of Allergies (2nd ed.) Facts on File, New York, p.167, ISBN0-8160-4404-X
Samuelsson, Bengt (ed.) (2001) Advances in prostaglandin and leukotriene research: basic science and new clinical applications: 11th International Conference on Advances in Prostaglandin and Leukotriene Research: Basic Science and New Clinical Applications, Florence, Italy, June 4–8, 2000 Kluwer Academic Publishers, Dordrecht, ISBN1-4020-0146-0
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.