| |
| Names | |
|---|---|
| Preferred IUPAC name 4-{(2S,3S)-3-[(1E,3E,5Z,8Z)-Tetradeca-1,3,5,8-tetraen-1-yl]oxiran-2-yl}butanoic acid | |
| Identifiers | |
3D model (JSmol) | |
| Abbreviations | LTA4 |
| ChEBI | |
| ChemSpider | |
| KEGG | |
| MeSH | D017572 |
PubChem CID | |
| UNII | |
CompTox Dashboard (EPA) | |
| |
| |
| Properties | |
| C20H30O3 | |
| Molar mass | 318.457 g·mol−1 |
Except where otherwise noted, data are given for materials in their standard state (at 25 °C [77 °F], 100 kPa). | |
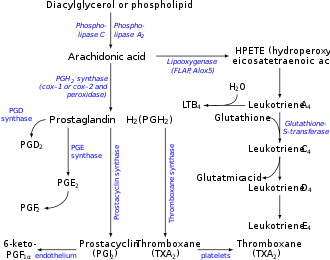
Leukotriene A4 (LTA4) is a leukotriene, and is the precursor for the productions of leukotriene B4 (LTB4) and leukotriene C4 (LTC4).