
Glycolysis is the metabolic pathway that converts glucose into pyruvate and, in most organisms, occurs in the liquid part of cells. The free energy released in this process is used to form the high-energy molecules adenosine triphosphate (ATP) and reduced nicotinamide adenine dinucleotide (NADH). Glycolysis is a sequence of ten reactions catalyzed by enzymes.

Xylitol is a chemical compound with the formula C
5H
12O
5, or HO(CH2)(CHOH)3(CH2)OH; specifically, one particular stereoisomer with that structural formula. It is a colorless or white crystalline solid that is freely soluble in water. It is classified as a polyalcohol and a sugar alcohol, specifically an alditol. The name derives from Ancient Greek: ξύλον, xyl[on] 'wood', with the suffix -itol used to denote it being a sugar alcohol.
In biochemistry, isomerases are a general class of enzymes that convert a molecule from one isomer to another. Isomerases facilitate intramolecular rearrangements in which bonds are broken and formed. The general form of such a reaction is as follows:
In organic chemistry, a tetrose is a monosaccharide with 4 carbon atoms. They have either an aldehyde functional group in position 1 (aldotetroses) or a ketone group in position 2 (ketotetroses).
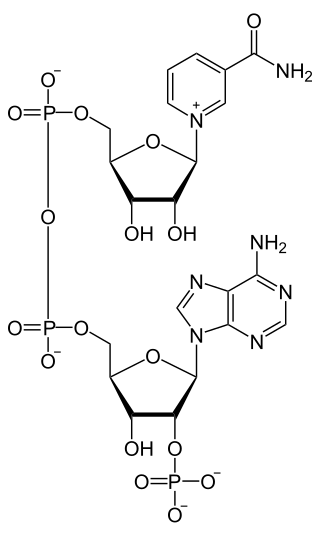
Nicotinamide adenine dinucleotide phosphate, abbreviated NADP or, in older notation, TPN (triphosphopyridine nucleotide), is a cofactor used in anabolic reactions, such as the Calvin cycle and lipid and nucleic acid syntheses, which require NADPH as a reducing agent ('hydrogen source'). NADPH is the reduced form, whereas NADP+ is the oxidized form. NADP+ is used by all forms of cellular life.
Cellulosic ethanol is ethanol produced from cellulose rather than from the plant's seeds or fruit. It can be produced from grasses, wood, algae, or other plants. It is generally discussed for use as a biofuel. The carbon dioxide that plants absorb as they grow offsets some of the carbon dioxide emitted when ethanol made from them is burned, so cellulosic ethanol fuel has the potential to have a lower carbon footprint than fossil fuels.

Kilju is the Finnish word for home made alcoholic beverage typically made of sugar, yeast, and water. The ABV is around 15–17%, and since it does not contain a sweet reserve it is completely dry. Crude fermented water may be distilled to moonshine. Kilju for consumption is clarified to avoid wine fault. It is a flax-colored alcoholic beverage with no discernible taste other than that of ethanol. It can be used as an ethanol base for drink mixers.
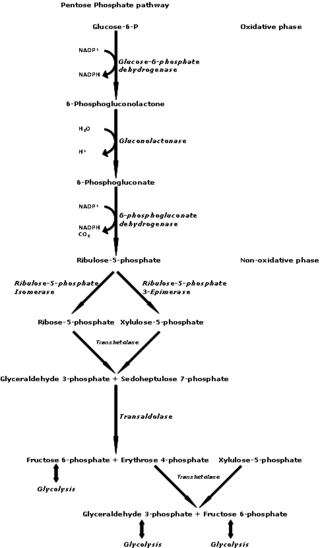
The pentose phosphate pathway is a metabolic pathway parallel to glycolysis. It generates NADPH and pentoses as well as ribose 5-phosphate, a precursor for the synthesis of nucleotides. While the pentose phosphate pathway does involve oxidation of glucose, its primary role is anabolic rather than catabolic. The pathway is especially important in red blood cells (erythrocytes). The reactions of the pathway were elucidated in the early 1950s by Bernard Horecker and co-workers.
Zymomonas mobilis is a Gram negative, facultative anaerobic, non-sporulating, polarly-flagellated, rod-shaped bacterium. It is the only species found in the genus Zymomonas. It has notable bioethanol-producing capabilities, which surpass yeast in some aspects. It was originally isolated from alcoholic beverages like the African palm wine, the Mexican pulque, and also as a contaminant of cider and beer in European countries.
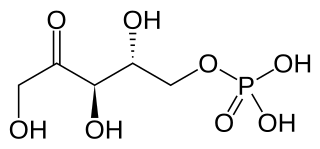
Ribulose 5-phosphate is one of the end-products of the pentose phosphate pathway. It is also an intermediate in the Calvin cycle.

Phosphopentose epimerase encoded in humans by the RPE gene is a metalloprotein that catalyzes the interconversion between D-ribulose 5-phosphate and D-xylulose 5-phosphate.

6-Phosphogluconolactonase (EC 3.1.1.31, 6PGL, PGLS, systematic name 6-phospho-D-glucono-1,5-lactone lactonohydrolase) is a cytosolic enzyme found in all organisms that catalyzes the hydrolysis of 6-phosphogluconolactone to 6-phosphogluconic acid in the oxidative phase of the pentose phosphate pathway:

In enzymology, a D-xylulose reductase (EC 1.1.1.9) is an enzyme that is classified as an Oxidoreductase (EC 1) specifically acting on the CH-OH group of donors (EC 1.1.1) that uses NAD+ or NADP+ as an acceptor (EC 1.1.1.9). This enzyme participates in pentose and glucuronate interconversions; a set of metabolic pathways that involve converting pentose sugars and glucuronate into other compounds.
In enzymology, a phosphogluconate dehydrogenase (decarboxylating) (EC 1.1.1.44) is an enzyme that catalyzes the chemical reaction
The enzyme phosphoketolase(EC 4.1.2.9) catalyzes the chemical reactions
Scheffersomyces stipitis is a species of yeast, belonging to the "CUG Clade" of ascomycetous yeasts. This is a group of fungi that substitute serine for leucine when the CUG codon is encountered. S. stipitis is distantly related to brewer's yeast, Saccharomyces cerevisiae, which uses the conventional codon system. Found, among other places, in the guts of passalid beetles, S. stipitis is capable of both aerobic and oxygen limited fermentation, and has the highest known natural ability of any yeast to directly ferment xylose, converting it to ethanol, a potentially economically valuable trait. Xylose is a hemicellulosic sugar found in all angiosperm plants. As such xylose constitutes the second most abundant carbohydrate moiety in nature. Xylose can be produced from wood or agricultural residues through auto- or acid hydrolysis. Ethanol production from such lignocellulosic residues does not compete with food production through the consumption of grain.
Cofactor engineering, a subset of metabolic engineering, is defined as the manipulation of the use of cofactors in an organism’s metabolic pathways. In cofactor engineering, the concentrations of cofactors are changed in order to maximize or minimize metabolic fluxes. This type of engineering can be used to optimize the production of a metabolite product or to increase the efficiency of a metabolic network. The use of engineering single celled organisms to create lucrative chemicals from cheap raw materials is growing, and cofactor engineering can play a crucial role in maximizing production. The field has gained more popularity in the past decade and has several practical applications in chemical manufacturing, bioengineering and pharmaceutical industries.
D-xylose reductase (EC 1.1.1.307, XylR, XyrA, msXR, dsXR, monospecific xylose reductase, dual specific xylose reductase, NAD(P)H-dependent xylose reductase, xylose reductase) is an enzyme with systematic name xylitol:NAD(P)+ oxidoreductase. This enzyme catalyses the following chemical reaction

In enzymology, a xylose isomerase is an enzyme that catalyzes the interconversion of D-xylose and D-xylulose. This enzyme belongs to the family of isomerases, specifically those intramolecular oxidoreductases interconverting aldoses and ketoses. The isomerase has now been observed in nearly a hundred species of bacteria. Xylose-isomerases are also commonly called fructose-isomerases due to their ability to interconvert glucose and fructose. The systematic name of this enzyme class is D-xylose aldose-ketose-isomerase. Other names in common use include D-xylose isomerase, D-xylose ketoisomerase, and D-xylose ketol-isomerase.
Aerobic fermentation or aerobic glycolysis is a metabolic process by which cells metabolize sugars via fermentation in the presence of oxygen and occurs through the repression of normal respiratory metabolism. Preference of aerobic fermentation over aerobic respiration is referred to as the Crabtree effect in yeast, and is part of the Warburg effect in tumor cells. While aerobic fermentation does not produce adenosine triphosphate (ATP) in high yield, it allows proliferating cells to convert nutrients such as glucose and glutamine more efficiently into biomass by avoiding unnecessary catabolic oxidation of such nutrients into carbon dioxide, preserving carbon-carbon bonds and promoting anabolism.
