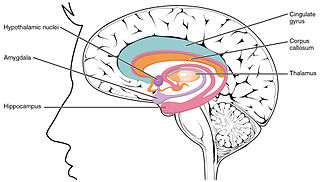
The hippocampus is a major component of the brain of humans and other vertebrates. Humans and other mammals have two hippocampi, one in each side of the brain. The hippocampus is part of the limbic system, and plays important roles in the consolidation of information from short-term memory to long-term memory, and in spatial memory that enables navigation. The hippocampus is located in the allocortex, with neural projections into the neocortex, in humans as well as other primates. The hippocampus, as the medial pallium, is a structure found in all vertebrates. In humans, it contains two main interlocking parts: the hippocampus proper, and the dentate gyrus.
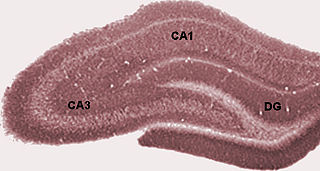
The dentate gyrus (DG) is part of the hippocampal formation in the temporal lobe of the brain, which also includes the hippocampus and the subiculum. The dentate gyrus is part of the hippocampal trisynaptic circuit and is thought to contribute to the formation of new episodic memories, the spontaneous exploration of novel environments and other functions.
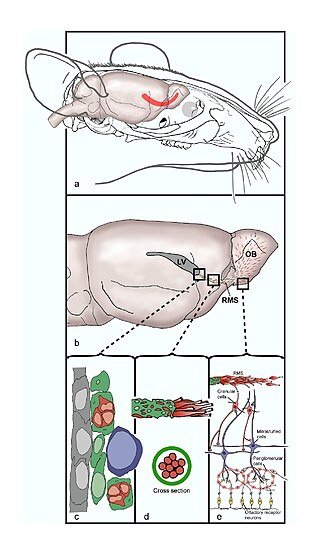
The rostral migratory stream (RMS) is a specialized migratory route found in the brain of some animals along which neuronal precursors that originated in the subventricular zone (SVZ) of the brain migrate to reach the main olfactory bulb (OB). The importance of the RMS lies in its ability to refine and even change an animal's sensitivity to smells, which explains its importance and larger size in the rodent brain as compared to the human brain, as our olfactory sense is not as developed. This pathway has been studied in the rodent, rabbit, and both the squirrel monkey and rhesus monkey. When the neurons reach the OB they differentiate into GABAergic interneurons as they are integrated into either the granule cell layer or periglomerular layer.
Elizabeth Gould is an American neuroscientist and the Dorman T. Warren Professor of Psychology at Princeton University. She was an early investigator of adult neurogenesis in the hippocampus, a research area that continues to be controversial. In November 2002, Discover magazine listed her as one of the 50 most important women scientists.
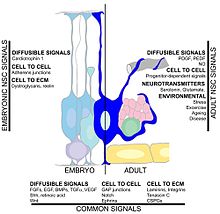
Neural stem cells (NSCs) are self-renewing, multipotent cells that firstly generate the radial glial progenitor cells that generate the neurons and glia of the nervous system of all animals during embryonic development. Some neural progenitor stem cells persist in highly restricted regions in the adult vertebrate brain and continue to produce neurons throughout life. Differences in the size of the central nervous system are among the most important distinctions between the species and thus mutations in the genes that regulate the size of the neural stem cell compartment are among the most important drivers of vertebrate evolution.
Neuropoiesis is the process by which neural stem cells differentiate to form mature neurons, astrocytes, and oligodendrocytes in the adult mammal. This process is also referred to as adult neurogenesis.

The subventricular zone (SVZ) is a region situated on the outside wall of each lateral ventricle of the vertebrate brain. It is present in both the embryonic and adult brain. In embryonic life, the SVZ refers to a secondary proliferative zone containing neural progenitor cells, which divide to produce neurons in the process of neurogenesis. The primary neural stem cells of the brain and spinal cord, termed radial glial cells, instead reside in the ventricular zone (VZ).

The subgranular zone (SGZ) is a brain region in the hippocampus where adult neurogenesis occurs. The other major site of adult neurogenesis is the subventricular zone (SVZ) in the brain.
Radiation-induced cognitive decline describes the possible correlation between radiation therapy and cognitive impairment. Radiation therapy is used mainly in the treatment of cancer. Radiation therapy can be used to cure care or shrink tumors that are interfering with quality of life. Sometimes radiation therapy is used alone; other times it is used in conjunction with chemotherapy and surgery. For people with brain tumors, radiation can be an effective treatment because chemotherapy is often less effective due to the blood–brain barrier. Unfortunately for some patients, as time passes, people who received radiation therapy may begin experiencing deficits in their learning, memory, and spatial information processing abilities. The learning, memory, and spatial information processing abilities are dependent on proper hippocampus functionality. Therefore, any hippocampus dysfunction will result in deficits in learning, memory, and spatial information processing ability.
Protein BTG2 also known as BTG family member 2 or NGF-inducible anti-proliferative protein PC3 or NGF-inducible protein TIS21, is a protein that in humans is encoded by the BTG2 gene and in other mammals by the homologous Btg2 gene. This protein controls cell cycle progression and proneural genes expression by acting as a transcription coregulator that enhances or inhibits the activity of transcription factors.
The trisynaptic circuit or trisynaptic loop is a relay of synaptic transmission in the hippocampus. The circuit was initially described by the neuroanatomist Santiago Ramon y Cajal, in the early twentieth century, using the Golgi staining method. After the discovery of the trisynaptic circuit, a series of research has been conducted to determine the mechanisms driving this circuit. Today, research is focused on how this loop interacts with other parts of the brain, and how it influences human physiology and behaviour. For example, it has been shown that disruptions within the trisynaptic circuit lead to behavioural changes in rodent and feline models.

Hippocampus anatomy describes the physical aspects and properties of the hippocampus, a neural structure in the medial temporal lobe of the brain. It has a distinctive, curved shape that has been likened to the sea-horse monster of Greek mythology and the ram's horns of Amun in Egyptian mythology. This general layout holds across the full range of mammalian species, from hedgehog to human, although the details vary. For example, in the rat, the two hippocampi look similar to a pair of bananas, joined at the stems. In primate brains, including humans, the portion of the hippocampus near the base of the temporal lobe is much broader than the part at the top. Due to the three-dimensional curvature of this structure, two-dimensional sections such as shown are commonly seen. Neuroimaging pictures can show a number of different shapes, depending on the angle and location of the cut.

The name granule cell has been used for a number of different types of neurons whose only common feature is that they all have very small cell bodies. Granule cells are found within the granular layer of the cerebellum, the dentate gyrus of the hippocampus, the superficial layer of the dorsal cochlear nucleus, the olfactory bulb, and the cerebral cortex.
Endogenous regeneration in the brain is the ability of cells to engage in the repair and regeneration process. While the brain has a limited capacity for regeneration, endogenous neural stem cells, as well as numerous pro-regenerative molecules, can participate in replacing and repairing damaged or diseased neurons and glial cells. Another benefit that can be achieved by using endogenous regeneration could be avoiding an immune response from the host.
Epigenetic regulation of neurogenesis is the role that epigenetics plays in the regulation of neurogenesis.
Neurogenesis is the process by which nervous system cells, the neurons, are produced by neural stem cells (NSCs). In short, it is brain growth in relation to its organization. This occurs in all species of animals except the porifera (sponges) and placozoans. Types of NSCs include neuroepithelial cells (NECs), radial glial cells (RGCs), basal progenitors (BPs), intermediate neuronal precursors (INPs), subventricular zone astrocytes, and subgranular zone radial astrocytes, among others.
Attila Losonczy is a Hungarian neuroscientist, Professor of Neuroscience at Columbia University Medical Center. Losonczy's main area of research is on the relationship between neural networks and behavior, specifically with regard to learning in the hippocampus.
TLQP-62 (amino acid 556-617) is a VGF-derived C-terminal peptide that was first discovered by Trani et al. TLQP-62 is derived from VGF precursor protein via proteolytic cleavage by prohormone convertases PC1/3 at the RPR555 site. TLQP-62 is named after its first four N-terminal amino acids and its peptide length.
Adult neurogenesis is the process in which new neurons are born and subsequently integrate into functional brain circuits after birth and into adulthood. Avian species including songbirds are among vertebrate species that demonstrate particularly robust adult neurogenesis throughout their telencephalon, in contrast with the more limited neurogenic potential that are observed in adult mammals after birth. Adult neurogenesis in songbirds is observed in brain circuits that underlie complex specialized behavior, including the song control system and the hippocampus. The degree of postnatal and adult neurogenesis in songbirds varies between species, shows sexual dimorphism, fluctuates seasonally, and depends on hormone levels, cell death rates, and social environment. The increased extent of adult neurogenesis in birds compared to other vertebrates, especially in circuits that underlie complex specialized behavior, makes birds an excellent animal model to study this process and its functionality. Methods used in research to track adult neurogenesis in birds include the use of thymidine analogues and identifying endogenous markers of neurogenesis. Historically, the discovery of adult neurogenesis in songbirds substantially contributed to establishing the presence of adult neurogenesis and to progressing a line of research tightly associated with many potential clinical applications.
Adult neurogenesis is the process by which functional, mature neurons are produced from neural stem cells (NSCs) in the adult brain. In most mammals, including humans, it only occurs in the subgranular zone of the hippocampus, and in the olfactory bulb. The neurogenesis hypothesis of depression proposes that major depressive disorder is caused, at least partly, by impaired neurogenesis in the subgranular zone of the hippocampus.