Isomerases are a general class of enzymes that convert a molecule from one isomer to another. Isomerases facilitate intramolecular rearrangements in which bonds are broken and formed. The general form of such a reaction is as follows:

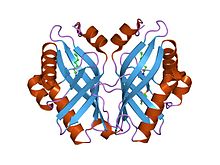
Aspartate transaminase (AST) or aspartate aminotransferase, also known as AspAT/ASAT/AAT or (serum) glutamic oxaloacetic transaminase, is a pyridoxal phosphate (PLP)-dependent transaminase enzyme that was first described by Arthur Karmen and colleagues in 1954. AST catalyzes the reversible transfer of an α-amino group between aspartate and glutamate and, as such, is an important enzyme in amino acid metabolism. AST is found in the liver, heart, skeletal muscle, kidneys, brain, red blood cells and gall bladder. Serum AST level, serum ALT level, and their ratio are commonly measured clinically as biomarkers for liver health. The tests are part of blood panels.

Epoxide hydrolases (EH's), also known as epoxide hydratases, are enzymes that metabolize compounds that contain an epoxide residue; they convert this residue to two hydroxyl residues through an epoxide hydrolysis reaction to form diol products. Several enzymes possess EH activity. Microsomal epoxide hydrolase, soluble epoxide hydrolase, and the more recently discovered but not as yet well defined functionally, epoxide hydrolase 3 (EH3) and epoxide hydrolase 4 (EH4) are structurally closely related isozymes. Other enzymes with epoxide hydrolase activity include leukotriene A4 hydrolase, Cholesterol-5,6-oxide hydrolase, MEST (gene) (Peg1/MEST), and Hepoxilin-epoxide hydrolase. The hydrolases are distinguished from each other by their substrate preferences and, directly related to this, their functions.
A halohydrin dehalogenase is an enzyme involved in the bacterial degradation of vicinal halohydrins. In several species of bacteria, it catalyses the dehalogenation of halohydrins to produce the corresponding epoxides. Different isoforms of the enzyme fall into one of three groups, A, B or C. Halogenases of the same class are genetically similar, but differ greatly from halogenases from a different group. Currently the most well-studied isoform is HheC which is purified from the bacterial species Agrobacterium radiobacter. The ability to dehalogenate organic compounds as well as form enantiomeric selective epoxides have generated interest in the potential of this enzyme in the biochemical field.

A catalytic triad is a set of three coordinated amino acids that can be found in the active site of some enzymes. Catalytic triads are most commonly found in hydrolase and transferase enzymes. An acid-base-nucleophile triad is a common motif for generating a nucleophilic residue for covalent catalysis. The residues form a charge-relay network to polarise and activate the nucleophile, which attacks the substrate, forming a covalent intermediate which is then hydrolysed to release the product and regenerate free enzyme. The nucleophile is most commonly a serine or cysteine amino acid, but occasionally threonine or even selenocysteine. The 3D structure of the enzyme brings together the triad residues in a precise orientation, even though they may be far apart in the sequence.

The enzyme citrate synthase E.C. 2.3.3.1 ] exists in nearly all living cells and stands as a pace-making enzyme in the first step of the citric acid cycle. Citrate synthase is localized within eukaryotic cells in the mitochondrial matrix, but is encoded by nuclear DNA rather than mitochondrial. It is synthesized using cytoplasmic ribosomes, then transported into the mitochondrial matrix.

Glycoside hydrolases catalyze the hydrolysis of glycosidic bonds in complex sugars. They are extremely common enzymes with roles in nature including degradation of biomass such as cellulose (cellulase), hemicellulose, and starch (amylase), in anti-bacterial defense strategies, in pathogenesis mechanisms and in normal cellular function. Together with glycosyltransferases, glycosidases form the major catalytic machinery for the synthesis and breakage of glycosidic bonds.

Sucrose phosphorylase is an important enzyme in the metabolism of sucrose and regulation of other metabolic intermediates. Sucrose phosphorylase is in the class of hexosyltransferases. More specifically it has been placed in the retaining glycoside hydrolases family although it catalyzes a transglycosidation rather than hydrolysis. Sucrose phosphorylase catalyzes the conversion of sucrose to D-fructose and α-D-glucose-1-phosphate. It has been shown in multiple experiments that the enzyme catalyzes this conversion by a double displacement mechanism.

In enzymology, a microsomal epoxide hydrolase (mEH) is an enzyme that catalyzes the hydrolysis reaction between an epoxide and water to form a diol.

The enzyme polyneuridine-aldehyde esterase (EC 3.1.1.78) catalyzes the following reaction:

In enzymology, an amidase is an enzyme that catalyzes the hydrolysis of an amide:
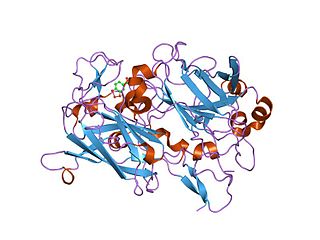
Dioxygenases are oxidoreductase enzymes. Aerobic life, from simple single-celled bacteria species to complex eukaryotic organisms, has evolved to depend on the oxidizing power of dioxygen in various metabolic pathways. From energetic adenosine triphosphate (ATP) generation to xenobiotic degradation, the use of dioxygen as a biological oxidant is widespread and varied in the exact mechanism of its use. Enzymes employ many different schemes to use dioxygen, and this largely depends on the substrate and reaction at hand.

The CFTR inhibitory factor (Cif) is a protein virulence factor secreted by the Gram-negative bacteria Pseudomonas aeruginosa and Acinetobacter nosocomialis. Discovered at Dartmouth Medical School, Cif is able to alter the trafficking of select ABC transporters in eukaryotic epithelial cells, such as the cystic fibrosis transmembrane conductance regulator (CFTR), and P-glycoprotein by interfering with the host deubiquitinating machinery. By promoting the ubiquitin-mediated degradation of CFTR, Cif is able to phenocopy cystic fibrosis at the cellular level. The cif gene is transcribed as part of a 3 gene operon, whose expression is negatively regulated by CifR, a TetR family repressor.
Dihydrocarveol dehydrogenase (EC 1.1.1.296, carveol dehydrogenase (ambiguous)) is an enzyme with systematic name menth-8-en-2-ol:NAD+ oxidoreductase. This enzyme catalyses the following chemical reaction
Limonene-1,2-diol dehydrogenase (EC 1.1.1.297, NAD+-dependent limonene-1,2-diol dehydrogenase) is an enzyme with systematic name menth-8-ene-1,2-diol:NAD+ oxidoreductase. This enzyme catalyses the following chemical reaction
Monocyclic monoterpene ketone monooxygenase (EC 1.14.13.105, 1-hydroxy-2-oxolimonene 1,2-monooxygenase, dihydrocarvone 1,2-monooxygenase, MMKMO) is an enzyme with systematic name (-)-menthone,NADPH:oxygen oxidoreductase. This enzyme catalyses the following chemical reaction
Limonene 1,2-monooxygenase (EC 1.14.13.107) is an enzyme with systematic name limonene,NAD(P)H:oxygen oxidoreductase. This enzyme catalyses the following chemical reaction
The enzyme monoterpene ε-lactone hydrolase (EC 3.1.1.83, MLH; systematic name isoprop(en)ylmethyloxepan-2-one lactonohydrolase catalyses the reaction

Neopullulanase is an enzyme of the alpha-amylase family with systematic name pullulan 4-D-glucanohydrolase (panose-forming). This enzyme principally catalyses the following chemical reaction by cleaving pullulan's alpha-1,4-glucosidic bonds:

Juvenile hormone epoxide hydrolase (JHEH) is an enzyme that inactivates insect juvenile hormones. This inactivation is accomplished through hydrolysis of the epoxide functional group contained within these hormones into diols. JHEH is one of two enzymes involved in the termination of signaling properties of the various juvenile hormones. The other is juvenile-hormone esterase, or JHE.