The inner ear is the innermost part of the vertebrate ear. In vertebrates, the inner ear is mainly responsible for sound detection and balance. In mammals, it consists of the bony labyrinth, a hollow cavity in the temporal bone of the skull with a system of passages comprising two main functional parts:

The cochlea is the part of the inner ear involved in hearing. It is a spiral-shaped cavity in the bony labyrinth, in humans making 2.75 turns around its axis, the modiolus. A core component of the cochlea is the organ of Corti, the sensory organ of hearing, which is distributed along the partition separating the fluid chambers in the coiled tapered tube of the cochlea.
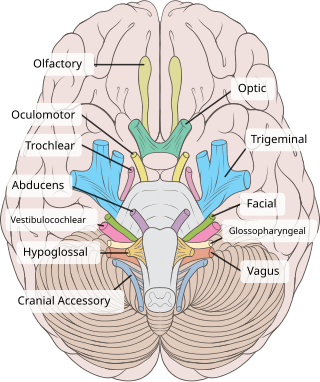
The vestibulocochlear nerve or auditory vestibular nerve, also known as the eighth cranial nerve, cranial nerve VIII, or simply CN VIII, is a cranial nerve that transmits sound and equilibrium (balance) information from the inner ear to the brain. Through olivocochlear fibers, it also transmits motor and modulatory information from the superior olivary complex in the brainstem to the cochlea.

The semicircular canals are three semicircular interconnected tubes located in the innermost part of each ear, the inner ear. The three canals are the lateral, anterior and posterior semicircular canals. They are the part of the bony labyrinth, a periosteum-lined cavity on the petrous part of the temporal bone filled with perilymph.

The utricle and saccule are the two otolith organs in the vertebrate inner ear. The word utricle comes from Latin uter 'leather bag'. The utricle and saccule are part of the balancing system in the vestibule of the bony labyrinth. They use small stones and a viscous fluid to stimulate hair cells to detect motion and orientation. The utricle detects linear accelerations and head-tilts in the horizontal plane.
The saccule is a bed of sensory cells in the inner ear that detects linear acceleration and head tilting in the vertical plane, and converts these vibrations into electrical impulses to be interpreted by the brain. When the head moves vertically, the sensory cells of the saccule are moved due to a combination of inertia and gravity. In response, the neurons connected to the saccule transmit electrical impulses that represent this movement to the brain. These impulses travel along the vestibular portion of the eighth cranial nerve to the vestibular nuclei in the brainstem.
The vestibular system, in vertebrates, is a sensory system that creates the sense of balance and spatial orientation for the purpose of coordinating movement with balance. Together with the cochlea, a part of the auditory system, it constitutes the labyrinth of the inner ear in most mammals.

The olfactory epithelium is a specialized epithelial tissue inside the nasal cavity that is involved in smell. In humans, it measures 5 cm2 (0.78 sq in) and lies on the roof of the nasal cavity about 7 cm (2.8 in) above and behind the nostrils. The olfactory epithelium is the part of the olfactory system directly responsible for detecting odors.

In vertebrates, an ear is the organ that enables hearing and body balance using the vestibular system. In humans, the ear is described as having three parts: the outer ear, the middle ear and the inner ear. The outer ear consists of the auricle and the ear canal. Since the outer ear is the only visible portion of the ear, the word "ear" often refers to the external part (auricle) alone. The middle ear includes the tympanic cavity and the three ossicles. The inner ear sits in the bony labyrinth, and contains structures which are key to several senses: the semicircular canals, which enable balance and eye tracking when moving; the utricle and saccule, which enable balance when stationary; and the cochlea, which enables hearing. The ear canal is cleaned via earwax, which naturally migrates to the auricle.
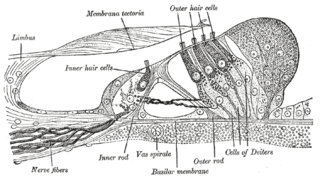
Hair cells are the sensory receptors of both the auditory system and the vestibular system in the ears of all vertebrates, and in the lateral line organ of fishes. Through mechanotransduction, hair cells detect movement in their environment.

In the inner ear, stereocilia are the mechanosensing organelles of hair cells, which respond to fluid motion in numerous types of animals for various functions, including hearing and balance. They are about 10–50 micrometers in length and share some similar features of microvilli. The hair cells turn the fluid pressure and other mechanical stimuli into electric stimuli via the many microvilli that make up stereocilia rods. Stereocilia exist in the auditory and vestibular systems.
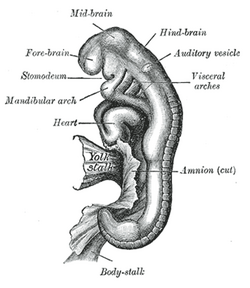
In embryology, a neurogenic placode is an area of thickening of the epithelium in the embryonic head ectoderm layer that gives rise to neurons and other structures of the sensory nervous system.
The vestibular ganglion is a collection of cell bodies belonging to first order sensory neurons of the vestibular nerve. It is located within the internal auditory canal.
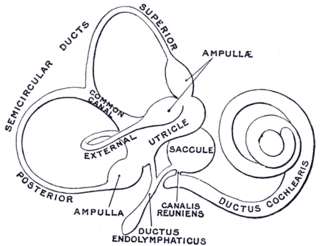
The membranous labyrinth is a collection of fluid filled tubes and chambers which contain the receptors for the senses of equilibrium and hearing. It is lodged within the bony labyrinth in the inner ear and has the same general form; it is, however, considerably smaller and is partly separated from the bony walls by a quantity of fluid, the perilymph.
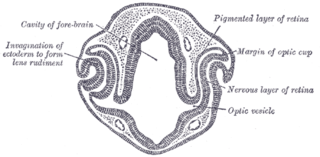
Eye formation in the human embryo begins at approximately three weeks into embryonic development and continues through the tenth week. Cells from both the mesodermal and the ectodermal tissues contribute to the formation of the eye. Specifically, the eye is derived from the neuroepithelium, surface ectoderm, and the extracellular mesenchyme which consists of both the neural crest and mesoderm.

The crista ampullaris is the sensory organ of rotation. They are found in the ampullae of each of the semicircular canals of the inner ear, meaning that there are three pairs in total. The function of the crista ampullaris is to sense angular acceleration and deceleration.
The vestibule is the central part of the bony labyrinth in the inner ear, and is situated medial to the eardrum, behind the cochlea, and in front of the three semicircular canals.
The vestibular evoked myogenic potential is a neurophysiological assessment technique used to determine the function of the otolithic organs of the inner ear. It complements the information provided by caloric testing and other forms of inner ear testing. There are two different types of VEMPs. One is the oVEMP and another is the cVEMP. The oVEMP measures integrity of the utricule and superior vestibular nerve and the cVemp measures the saccule and the inferior vestibular nerve.
Dark cells are specialized nonsensory epithelial cells found on either side of the vestibular organs and lining the endolymphatic space. These dark-cell areas in the vestibular organ are structures involved in the production of endolymph, an inner ear fluid, secreting potassium towards the endolymphatic fluid. Dark cells take part in fluid homeostasis to preserve the unique high-potassium and low-sodium content of the endolymph and also maintain the calcium homeostasis of the inner ear.

Otoconin is a structural glycoprotein found in the otoconial membrane of vertebrates. It is the major protein component of the otoconia.