
An echinoderm is any animal of the phylum Echinodermata, which includes starfish, brittle stars, sea urchins, sand dollars and sea cucumbers, as well as the sessile sea lilies or "stone lilies". While bilaterally symmetrical as larvae, as adults echinoderms are recognisable by their usually five-pointed radial symmetry, and are found on the sea bed at every ocean depth from the intertidal zone to the abyssal zone. The phylum contains about 7,600 living species, making it the second-largest group of deuterostomes after the chordates, as well as the largest marine-only phylum. The first definitive echinoderms appeared near the start of the Cambrian.

Helicoplacus is the earliest well-studied fossil echinoderm. Fossil plates are known from several regions. Complete specimens were found in Lower Cambrian strata of the White Mountains of California.

Cothurnocystis is a genus of small enigmatic echinoderms that lived during the Ordovician. Individual animals had a flat boot-shaped body and a thin rod-shaped appendage that may be a stem, or analogous to a foot or a tail. Fossils of Cothurnocystis species have been found in Nevada, Scotland, Czech Republic, France and Morocco.

Edrioasteroidea is an extinct class of echinoderms. The living animal would have resembled a pentamerously symmetrical disc or cushion. They were obligate encrusters and attached themselves to inorganic or biologic hard substrates. A 507 million years old species, Totiglobus spencensis, is actually the first known echinoderm adapted to live on a hard surface after the soft microbial mats that covered the seafloor were destroyed in the Cambrian substrate revolution.
The stylophorans are an extinct, possibly polyphyletic group allied to the Paleozoic Era echinoderms, comprising the prehistoric cornutes and mitrates. It is synonymous with the subphylum Calcichordata. Their unusual appearances have led to a variety of very different reconstructions of their anatomy, how they lived, and their relationships to other organisms.

Crinozoa is a subphylum of mostly sessile echinoderms, of which the crinoids, or sea lilies and feather stars, are the only extant members. Crinozoans have an extremely extensive fossil history.

Eleutherozoa is a subphylum of echinoderms. They are mobile animals with the mouth directed towards the substrate. They usually have a madreporite, tube feet, and moveable spines of some sort. It includes all living echinoderms except for crinoids. The monophyly of Eleutherozoa has been proven sufficiently well to be considered "uncontroversial."

The Eocrinoidea were an extinct class of echinoderms that lived between the Early Cambrian and Late Silurian periods. They are the earliest known group of stalked, brachiole-bearing echinoderms, and were the most common echinoderms during the Cambrian.

Camptostroma roddyi is an extinct echinoderm from the Bonnia-Olenellus Zone of the Early Cambrian Kinzers Formation near York and Lancaster, Southeastern Pennsylvania. It is the only known species in the genus Camptostroma, as other species referred to this genus "do not appear to be cogeneric."
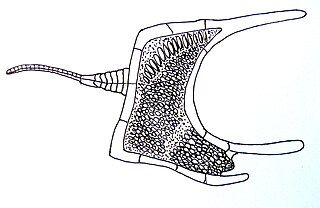
Homalozoa is an obsolete extinct subphylum of Paleozoic era echinoderms, prehistoric marine invertebrates. They are also referred to as carpoids.

Ophiocistioidea is a class of extinct echinoderms from the Palaeozoic and early Mesozoic. They most likely form a paraphyletic grade along sea cucumber stem lineage, although some sources still consider the question of ophiocistioid monophyly unresolved.
The calcichordate hypothesis, formulated by British Museum paleontologist Richard Jefferies, holds that each separate lineage of chordate evolved from its own lineage of mitrate, and thus the echinoderms and the chordates are sister groups, with the hemichordates as an out-group. It has been disproven by the discovery that the "tail" of Stylophorans contains a water vascular system, ambulacrum, and tube feet. However, the clade Olfactores was first proposed as part of the calcichordate theory, and has since been validated through genetic sequencing, albeit without the involvement of mitrates.

Stereom is a calcium carbonate material that makes up the internal skeletons found in all echinoderms, both living and fossilized forms. It is a sponge-like porous structure which, in a sea urchin may be 50% by volume living cells, and the rest being a matrix of calcite crystals. The size of openings in stereom varies in different species and in different places within the same organism. When an echinoderm becomes a fossil, microscopic examination is used to reveal the structure and such examination is often an important tool to classify the fossil as an echinoderm or related creature.
Diploporita is an extinct group of blastozoans that ranged from the Ordovician to the Devonian. These echinoderms are identified by a specialized respiratory structure, called diplopores. Diplopores are a double pore system that sit within a depression on a single thecal (body) plate; each plate can contain numerous diplopore pairs.

Soluta is an extinct class of echinoderms that lived from the Middle Cambrian to the Early Devonian. The class is also known by its junior synonym Homoiostelea. Soluta is one of the four "carpoid" classes, alongside Ctenocystoidea, Cincta, and Stylophora, which made up the obsolete subphylum Homalozoa. Solutes were asymmetric animals with a stereom skeleton and two appendages, an arm extending anteriorly and a posterior appendage called a homoiostele.

Ctenocystoidea is an extinct clade of echinoderms, which lived during the Cambrian and Ordovician periods. Unlike other echinoderms, ctenocystoids had bilateral symmetry, or were only very slightly asymmetrical. They are believed to be one of the earliest-diverging branches of echinoderms, with their bilateral symmetry a trait shared with other deuterostomes. Ctenocystoids were once classified in the taxon Homalozoa, also known as Carpoidea, alongside cinctans, solutes, and stylophorans. Homalozoa is now recognized as a polyphyletic group of echinoderms without radial symmetry. Ctenocystoids were geographically widespread during the Middle Cambrian, with one species surviving into the Late Ordovician.

Yanjiahella biscarpa is an extinct species of Early Cambrian deuterostome which may represent the earliest stem group echinoderm.

Vetulocystidae is the only family of the taxon Vetulocystida, which is a group of extinct deuterostomes of uncertain phylogenetic position. Vetulocystidae is made up of the genera Vetulocystis, Dianchicystis and Thylacocercus.

Lepidocystis is a Palaeozoic genus of imbricate lepidocystoid eocrinoid, closely related to Kinzercystis, It bore a stalk, with which it attached to firm substrates; and brachioles arising as lateral branches from its arms.

Protocinctus is a genus of cinctan and, as of 2024, it is the oldest such recognized genus. It is known from the Wuliuan age of the Mansilla Formation, Zaragoza, Spain. It is represented by the single species Protocinctus mansillaensis.
