
Cecropia is a Neotropical genus consisting of 61 recognized species with a highly distinctive lineage of dioecious trees. The genus consists of pioneer trees in the more or less humid parts of the Neotropics, with the majority of the species being myrmecophytic. Berg and Rosselli state that the genus is characterized by some unusual traits: spathes fully enclosing the flower-bearing parts of the inflorescences until anthesis, patches of dense indumentums (trichilia) producing Mullerian (food) at the base of the petiole, and anthers becoming detached at anthesis. Cecropia is most studied for its ecological role and association with ants. Its classification is controversial; in the past, it has been placed in the Cecropiaceae, Moraceae, or Urticaceae. The modern Angiosperm Phylogeny Group system places the "cecropiacean" group in the Urticaceae.

Calamagrostis is a genus of flowering plants in the grass family Poaceae, with about 260 species that occur mainly in temperate regions of the globe. Towards equatorial latitudes, species of Calamagrostis generally occur at higher elevations. These tufted perennials usually have hairless narrow leaves. The ligules are usually blunt. The inflorescence forms a panicle. Some may be reed-like.

Aiphanes is a genus of spiny palms which is native to tropical regions of South and Central America and the Caribbean. There are about 26 species in the genus, ranging in size from understorey shrubs with subterranean stems to subcanopy trees as tall as 20 metres (66 ft). Most have pinnately compound leaves ; one species has entire leaves. Stems, leaves and sometimes even the fruit are covered with spines. Plants flower repeatedly over the course of their lifespan and have separate male and female flowers, although these are borne together on the same inflorescence. Although records of pollinators are limited, most species appear to be pollinated by insects. The fruit are eaten by several birds and mammals, including at least two species of amazon parrots.

Fraxinus latifolia, the Oregon ash, is a member of the ash genus Fraxinus, native to western North America.
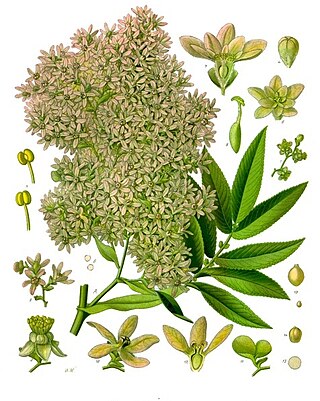
Hagenia is a monotypic genus of flowering plant with the sole species Hagenia abyssinica, native to the high-elevation Afromontane regions of central and eastern Africa. It also has a disjunct distribution in the high mountains of East Africa from Sudan and Ethiopia in the north, through Kenya, Uganda, Rwanda, Burundi, Democratic Republic of Congo, and Tanzania, to Malawi and Zambia in the south. A member of the rose family, its closest relative is the Afromontane genus Leucosidea.

Oemleria cerasiformis, a shrub commonly known as osoberry, squaw plum, Indian plum, or Indian Peach, is the sole extant species in genus Oemleria. The deciduous and perennial osoberry is one of the first plants to have its flowers bloom and leaves bud in the late winter. The shrub can grow up to 7 meters tall, with spread out branches that grow small white flowers and bitter fruit that sweeten when ripened. Osoberry is dioecious, with females producing fruit, and males producing a high reproductive biomass of pollen and flowers. Animals and insects aid in internal animal dispersion of seeds and pollination.

Polylepis australis, also known locally as tabaquillo or queñoa is a tree endemic of central Argentina, member of the family Rosaceae. The genus Polylepis originated in the eastern Andean forests of eastern South America.
This page provides a glossary of plant morphology. Botanists and other biologists who study plant morphology use a number of different terms to classify and identify plant organs and parts that can be observed using no more than a handheld magnifying lens. This page provides help in understanding the numerous other pages describing plants by their various taxa. The accompanying page—Plant morphology—provides an overview of the science of the external form of plants. There is also an alphabetical list: Glossary of botanical terms. In contrast, this page deals with botanical terms in a systematic manner, with some illustrations, and organized by plant anatomy and function in plant physiology.

Aloe comosa is a species of flowering plant in the Asphodelaceae family. It is commonly called Clanwilliam aloe) and is endemic to South Africa.

Polylepis racemosa is a species of small tree in the family Rosaceae. It is endemic to Peru, Bolivia, and Ecuador. It is threatened by habitat destruction. The International Union for Conservation of Nature has assessed the conservation status of this tree as "vulnerable".

Polylepis rugulosa, the queñua, is a species of plant in the family Rosaceae. It is primarily found in the Andes region of South America in Argentina, Bolivia, Chile and Peru. It is currently threatened by habitat loss.

Hedlundia hybrida, the Swedish service-treeFinnish whitebeam, or oakleaf mountain ash, is a species of whitebeam native to Norway, eastern Sweden, south-western Finland, and locally in Latvia.

Aphandra is a monotypic genus of flowering plants in the palm family native to the Amazon rainforest vegetation in South America. Its only species is Aphandra natalia, sometimes called mastodon palm or fiber palm, and is used by indigenous peoples in the construction of brooms and other products. This plant is commercially exploited for its edible fruits, and for its leaf sheath and petiole fibers. This fiber is almost equal to the fiber extracted from Attalea funifera and Leopoldinia piassaba, which is called piassava.
This glossary of botanical terms is a list of definitions of terms and concepts relevant to botany and plants in general. Terms of plant morphology are included here as well as at the more specific Glossary of plant morphology and Glossary of leaf morphology. For other related terms, see Glossary of phytopathology, Glossary of lichen terms, and List of Latin and Greek words commonly used in systematic names.

Lyonothamnus is a monotypic genus of trees in the rose family containing the single living species Lyonothamnus floribundus, which is known by the common name Catalina ironwood, and the subspecies L. f. ssp. aspleniifolius and L. f. ssp. floribundus.
Aiphanes deltoidea is a species of palm which is native to northeastern South America.

Rubus flagellaris, the northern dewberry, also known as the common dewberry, is a North American species perennial subshrub species of dewberry, in the rose family. This dewberry is distributed across much of Canada, Mexico, and the United States. It grows in diverse habitats ranging from drier savannas to temperate deciduous forests.

Cliffortia, or Caperose is a genus of plants that has been assigned to the rose family, with currently 132 known species. Its species can be found in southern Africa, particularly in the Cape Floristic Region where 124 of the species can be found, 109 of which are endemic to the CFR. Most species are ericoid shrubs, some small trees up to 5 m high, others more or less herbaceous groundcover. All are wind pollinated and have separate male and female flowers in the axils of the leaves, mostly individually, sometimes grouped, which may be on the same plant or on separate plants.
Acaena alpina is a perennial shrub of the genus Acaena known for its hardiness and durability. A. alpina is found throughout central Chile and Argentina. It can withstand a wide range of climates, including that of the Andes, where it is commonly found. A.alpina can withstand both hot and cold temperatures as well as wet and dry seasons, though it preferentially grows at high altitudes. A. alpina was originally typified by Eduard Friedrich Poeppig and Wilhelm Gerhard Walpers in 1843.

Ourisia chamaedrifolia is a species of flowering plant in the family Plantaginaceae that is endemic to páramo habitats in the Tropical Andes mountains of Venezuela, Colombia, Ecuador, Peru and Bolivia. George Bentham described O. chamaedrifolia in 1846. Plants of this species of South American foxglove are small, perennial, and repent herbs with opposite, crenate, and often hairy leaves. There can be up to four flowers on a short raceme, and each flower has a regular calyx, and a long, tubular, red or orange-red nearly bilabiate corolla with exserted stamens. The calyx and corolla are often hairless.

