
Symbiosis is any type of a close and long-term biological interaction between two biological organisms of different species, termed symbionts, be it mutualistic, commensalistic, or parasitic. In 1879, Heinrich Anton de Bary defined it as "the living together of unlike organisms". The term is sometimes used in the more restricted sense of a mutually beneficial interaction in which both symbionts contribute to each other's support.

Mutualism describes the ecological interaction between two or more species where each species has a net benefit. Mutualism is a common type of ecological interaction, one that can come from a parasitic interaction. Prominent examples include most vascular plants engaged in mutualistic interactions with mycorrhizae, flowering plants being pollinated by animals, vascular plants being dispersed by animals, and corals with zooxanthellae, among many others. Mutualism can be contrasted with interspecific competition, in which each species experiences reduced fitness, and exploitation, or parasitism, in which one species benefits at the expense of the other.

A pollinator is an animal that moves pollen from the male anther of a flower to the female stigma of a flower. This helps to bring about fertilization of the ovules in the flower by the male gametes from the pollen grains.

In biology, coevolution occurs when two or more species reciprocally affect each other's evolution through the process of natural selection. The term sometimes is used for two traits in the same species affecting each other's evolution, as well as gene-culture coevolution.
Conservation biology is the study of the conservation of nature and of Earth's biodiversity with the aim of protecting species, their habitats, and ecosystems from excessive rates of extinction and the erosion of biotic interactions. It is an interdisciplinary subject drawing on natural and social sciences, and the practice of natural resource management.

In ecology, a biological interaction is the effect that a pair of organisms living together in a community have on each other. They can be either of the same species, or of different species. These effects may be short-term, or long-term, both often strongly influence the adaptation and evolution of the species involved. Biological interactions range from mutualism, beneficial to both partners, to competition, harmful to both partners. Interactions can be direct when physical contact is established or indirect, through intermediaries such as shared resources, territories, ecological services, metabolic waste, toxins or growth inhibitors. This type of relationship can be shown by net effect based on individual effects on both organisms arising out of relationship.

Habitat conservation is a management practice that seeks to conserve, protect and restore habitats and prevent species extinction, fragmentation or reduction in range. It is a priority of many groups that cannot be easily characterized in terms of any one ideology.
Coextinction and cothreatened refer to the phenomena of the loss or decline of a host species resulting in the loss or endangerment of an other species that depends on it, potentially leading to cascading effects across trophic levels. The term originated by the authors Stork and Lyal (1993) and was originally used to explain the extinction of parasitic insects following the loss of their specific hosts. The term is now used to describe the loss of any interacting species, including competition with their counterpart, and specialist herbivores with their food source. Coextinction is especially common when a keystone species goes extinct.
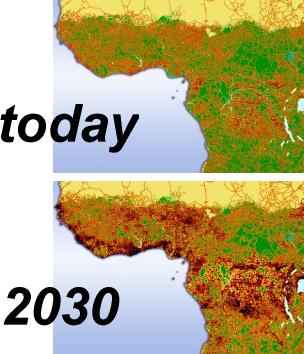
Habitat fragmentation describes the emergence of discontinuities (fragmentation) in an organism's preferred environment (habitat), causing population fragmentation and ecosystem decay. Causes of habitat fragmentation include geological processes that slowly alter the layout of the physical environment, and human activity such as land conversion, which can alter the environment much faster and causes the extinction of many species. More specifically, habitat fragmentation is a process by which large and contiguous habitats get divided into smaller, isolated patches of habitats.
Evolutionary ecology lies at the intersection of ecology and evolutionary biology. It approaches the study of ecology in a way that explicitly considers the evolutionary histories of species and the interactions between them. Conversely, it can be seen as an approach to the study of evolution that incorporates an understanding of the interactions between the species under consideration. The main subfields of evolutionary ecology are life history evolution, sociobiology, the evolution of interspecific interactions and the evolution of biodiversity and of ecological communities.
The Prodoxidae are a family of moths, generally small in size and nondescript in appearance. They include species of moderate pest status, such as the currant shoot borer, and others of considerable ecological and evolutionary interest, such as various species of "yucca moths".
Cheating is a term used in behavioral ecology and ethology to describe behavior whereby organisms receive a benefit at the cost of other organisms. Cheating is common in many mutualistic and altruistic relationships. A cheater is an individual who does not cooperate but can potentially gain the benefit from others cooperating. Cheaters are also those who selfishly use common resources to maximize their individual fitness at the expense of a group. Natural selection favors cheating, but there are mechanisms to regulate it. The stress gradient hypothesis states that facilitation, cooperation or mutualism should be more common in stressful environments, while cheating, competition or parasitism are common in benign environments.
An ecological cascade effect is a series of secondary extinctions that are triggered by the primary extinction of a key species in an ecosystem. Secondary extinctions are likely to occur when the threatened species are: dependent on a few specific food sources, mutualistic, or forced to coexist with an invasive species that is introduced to the ecosystem. Species introductions to a foreign ecosystem can often devastate entire communities, and even entire ecosystems. These exotic species monopolize the ecosystem's resources, and since they have no natural predators to decrease their growth, they are able to increase indefinitely. Olsen et al. showed that exotic species have caused lake and estuary ecosystems to go through cascade effects due to loss of algae, crayfish, mollusks, fish, amphibians, and birds. However, the principal cause of cascade effects is the loss of top predators as the key species. As a result of this loss, a dramatic increase of prey species occurs. The prey is then able to overexploit its own food resources, until the population numbers decrease in abundance, which can lead to extinction. When the prey's food resources disappear, they starve and may go extinct as well. If the prey species is herbivorous, then their initial release and exploitation of the plants may result in a loss of plant biodiversity in the area. If other organisms in the ecosystem also depend upon these plants as food resources, then these species may go extinct as well. An example of the cascade effect caused by the loss of a top predator is apparent in tropical forests. When hunters cause local extinctions of top predators, the predators' prey's population numbers increase, causing an overexploitation of a food resource and a cascade effect of species loss. Recent studies have been performed on approaches to mitigate extinction cascades in food-web networks.
Ecological extinction is "the reduction of a species to such low abundance that, although it is still present in the community, it no longer interacts significantly with other species".

A large proportion of living species on Earth live a parasitic way of life. Parasites have traditionally been seen as targets of eradication efforts, and they have often been overlooked in conservation efforts. In the case of parasites living in the wild – and thus harmless to humans and domesticated animals – this view is changing. The conservation biology of parasites is an emerging and interdisciplinary field that recognizes the integral role parasites play in ecosystems. Parasites are intricately woven into the fabric of ecological communities, with diverse species occupying a range of ecological niches and displaying complex relationships with their hosts.

Evolving digital ecological networks are webs of interacting, self-replicating, and evolving computer programs that experience the same major ecological interactions as biological organisms. Despite being computational, these programs evolve quickly in an open-ended way, and starting from only one or two ancestral organisms, the formation of ecological networks can be observed in real-time by tracking interactions between the constantly evolving organism phenotypes. These phenotypes may be defined by combinations of logical computations that digital organisms perform and by expressed behaviors that have evolved. The types and outcomes of interactions between phenotypes are determined by task overlap for logic-defined phenotypes and by responses to encounters in the case of behavioral phenotypes. Biologists use these evolving networks to study active and fundamental topics within evolutionary ecology.

A pollination network is a bipartite mutualistic network in which plants and pollinators are the nodes, and the pollination interactions form the links between these nodes. The pollination network is bipartite as interactions only exist between two distinct, non-overlapping sets of species, but not within the set: a pollinator can never be pollinated, unlike in a predator-prey network where a predator can be depredated. A pollination network is two-modal, i.e., it includes only links connecting plant and animal communities.
Colpocephalum californici, the California condor louse, is an extinct species of chewing louse which parasitized the California condor. In an example of coextinction, it became extinct when the remaining, Critically Endangered California condors were deloused and treated with pesticides during a captive breeding program.

Anna Traveset is a Spanish ecologist, particularly known for her work on ecological interactions between plants and animals, especially on islands.
Jordi Bascompte is a professor of ecology at the University of Zurich and the director of its specialized master's program on quantitative environmental sciences. He is best known for having brought the interactions of mutual benefit between plants and animals into community ecology, at the time largely dominated by predation and competition. His application of network theory to the study of mutualism has identified general laws that determine the way in which species interactions shape biodiversity.
