Extinction threshold is a term used in conservation biology to explain the point at which a species, population or metapopulation, experiences an abrupt change in density or number because of an important parameter, such as habitat loss. It is at this critical value below which a species, population, or metapopulation, will go extinct,[1] though this may take a long time for species just below the critical value, a phenomenon known as extinction debt.[2]
Extinction thresholds are important to conservation biologists when studying a species in a population or metapopulation context because the colonization rate must be larger than the extinction rate, otherwise the entire entity will go extinct once it reaches the threshold.[3]
Extinction thresholds are realized under a number of circumstances and the point in modeling them is to define the conditions that lead a population to extinction.[4] Modeling extinction thresholds can explain the relationship between extinction threshold and habitat loss and habitat fragmentation.[5]
Mathematical models
Metapopulation-type models are used to predict extinction thresholds. The classic metapopulation model is the Levins Model, which is the model of metapopulation dynamics established by Richard Levins in the 1960s. It was used to evaluate patch occupancy in a large network of patches. This model was extended in the 1980s by Russell Lande to include habitat occupancy.[1] This mathematical model is used to infer the extinction values and important population densities. These mathematical models are primarily used to study extinction thresholds because of the difficulty in understanding extinction processes through empirical methods and the current lack of research on this subject.[6] When determining an extinction threshold there are two types of models that can be used: deterministic and stochastic metapopulation models.
Deterministic
Deterministic metapopulation models assume that there are an infinite number of habitat patches available and predict that the metapopulation will go extinct only if the threshold is not met.[1]
dp/dt = chp (1-p)-ep
Where p= occupied patches, e= extinction rate, c= colonization rate, and h= amount of habitat.
A species will persist only if h> δ
where δ=e/c
δ= species parameter, or how successful a species is in colonizing a patch.[1]
Stochastic
Stochastic metapopulation models take into account stochasticity, which is the non-deterministic or random processes in nature. With this approach a metapopulation may be above the threshold if determined that it is unlikely it will go extinct within a certain time period.[1]
The complex nature of these models can result in a small metapopulation that is considered to be above the deterministic extinction threshold, but in reality has a high risk of extinction.[1]
Other factors
When using metapopulation-type models to predict extinction thresholds there are a number of factors that can affect the results of a model. First, including more complicated models, rather than relying solely on the Levins model produces different dynamics. For example, in an article published in 2004, Otso Ovaskainen and Ilkka Hanski explained with an empirical example that when factors such as Allee effect or Rescue effect were included in modeling the extinction threshold, there were unexpected extinctions in a high number of species. A more complex model came up with different results, and in practicing conservation biology this can add more confusion to efforts to save a species from the extinction threshold. Transient dynamics, which are effects on the extinction threshold because of instability in either the metapopulation or environmental conditions, is also a large player in modeling results. Landscapes that have recently endured habitat loss and fragmentation may be less able to sustain a metapopulation than previously understood without considering transient dynamics. Finally, environmental stochasticity, which may be spatially correlated, can lead to amplified regional stochastic fluctuations and therefore greatly affect the extinction risk.[1]
Theoretical ecology is the scientific discipline devoted to the study of ecological systems using theoretical methods such as simple conceptual models, mathematical models, computational simulations, and advanced data analysis. Effective models improve understanding of the natural world by revealing how the dynamics of species populations are often based on fundamental biological conditions and processes. Further, the field aims to unify a diverse range of empirical observations by assuming that common, mechanistic processes generate observable phenomena across species and ecological environments. Based on biologically realistic assumptions, theoretical ecologists are able to uncover novel, non-intuitive insights about natural processes. Theoretical results are often verified by empirical and observational studies, revealing the power of theoretical methods in both predicting and understanding the noisy, diverse biological world.
Population viability analysis (PVA) is a species-specific method of risk assessment frequently used in conservation biology. It is traditionally defined as the process that determines the probability that a population will go extinct within a given number of years. More recently, PVA has been described as a marriage of ecology and statistics that brings together species characteristics and environmental variability to forecast population health and extinction risk. Each PVA is individually developed for a target population or species, and consequently, each PVA is unique. The larger goal in mind when conducting a PVA is to ensure that the population of a species is self-sustaining over the long term.
Population ecology is a sub-field of ecology that deals with the dynamics of species populations and how these populations interact with the environment, such as birth and death rates, and by immigration and emigration.
Habitat fragmentation describes the emergence of discontinuities (fragmentation) in an organism's preferred environment (habitat), causing population fragmentation and ecosystem decay. Causes of habitat fragmentation include geological processes that slowly alter the layout of the physical environment, and human activity such as land conversion, which can alter the environment much faster and causes the extinction of many species. More specifically, habitat fragmentation is a process by which large and contiguous habitats get divided into smaller, isolated patches of habitats.
A metapopulation consists of a group of spatially separated populations of the same species which interact at some level. The term metapopulation was coined by Richard Levins in 1969 to describe a model of population dynamics of insect pests in agricultural fields, but the idea has been most broadly applied to species in naturally or artificially fragmented habitats. In Levins' own words, it consists of "a population of populations".
Insular biogeography or island biogeography is a field within biogeography that examines the factors that affect the species richness and diversification of isolated natural communities. The theory was originally developed to explain the pattern of the species–area relationship occurring in oceanic islands. Under either name it is now used in reference to any ecosystem that is isolated due to being surrounded by unlike ecosystems, and has been extended to mountain peaks, seamounts, oases, fragmented forests, and even natural habitats isolated by human land development. The field was started in the 1960s by the ecologists Robert H. MacArthur and E. O. Wilson, who coined the term island biogeography in their inaugural contribution to Princeton's Monograph in Population Biology series, which attempted to predict the number of species that would exist on a newly created island.
The Allee effect is a phenomenon in biology characterized by a correlation between population size or density and the mean individual fitness of a population or species.
Minimum viable population (MVP) is a lower bound on the population of a species, such that it can survive in the wild. This term is commonly used in the fields of biology, ecology, and conservation biology. MVP refers to the smallest possible size at which a biological population can exist without facing extinction from natural disasters or demographic, environmental, or genetic stochasticity. The term "population" is defined as a group of interbreeding individuals in similar geographic area that undergo negligible gene flow with other groups of the species. Typically, MVP is used to refer to a wild population, but can also be used for ex situ conservation.
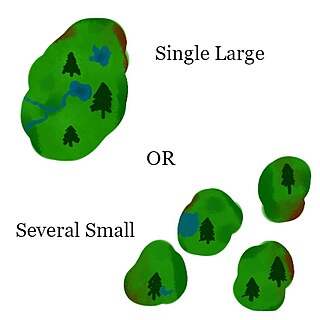
The SLOSS debate was a debate in ecology and conservation biology during the 1970's and 1980's as to whether a single large or several small (SLOSS) reserves were a superior means of conserving biodiversity in a fragmented habitat. Since its inception, multiple alternate theories have been proposed. There have been applications of the concept outside of the original context of habitat conservation.
Richard Levins was a Marxist biologist, a population geneticist, biomathematician, mathematical ecologist, and philosopher of science who researched diversity in human populations. Until his death, Levins was a university professor at the Harvard T.H. Chan School of Public Health and a long-time political activist. He was best known for his work on evolution and complexity in changing environments and on metapopulations.
In landscape ecology, landscape connectivity is, broadly, "the degree to which the landscape facilitates or impedes movement among resource patches". Alternatively, connectivity may be a continuous property of the landscape and independent of patches and paths. Connectivity includes both structural connectivity and functional connectivity. Functional connectivity includes actual connectivity and potential connectivity in which movement paths are estimated using the life-history data.
Ilkka Aulis Hanski was a Finnish ecologist at the University of Helsinki, Finland. The Metapopulation Research Center led by Hanski, until his death, has been nominated as a Center of Excellence by the Academy of Finland. The group studies species living in fragmented landscapes and attempts to advance metapopulation ecology research. Hanski proposed the core-satellite hypothesis of species distributions. Metapopulation ecology itself studies populations of plants and animals which are separated in space by occupying patches.
Source–sink dynamics is a theoretical model used by ecologists to describe how variation in habitat quality may affect the population growth or decline of organisms.
Patch dynamics is an ecological perspective that the structure, function, and dynamics of ecological systems can be understood through studying their interactive patches. Patch dynamics, as a term, may also refer to the spatiotemporal changes within and among patches that make up a landscape. Patch dynamics is ubiquitous in terrestrial and aquatic systems across organizational levels and spatial scales. From a patch dynamics perspective, populations, communities, ecosystems, and landscapes may all be studied effectively as mosaics of patches that differ in size, shape, composition, history, and boundary characteristics.
In ecology, the occupancy–abundance (O–A) relationship is the relationship between the abundance of species and the size of their ranges within a region. This relationship is perhaps one of the most well-documented relationships in macroecology, and applies both intra- and interspecifically. In most cases, the O–A relationship is a positive relationship. Although an O–A relationship would be expected, given that a species colonizing a region must pass through the origin and could reach some theoretical maximum abundance and distribution, the relationship described here is somewhat more substantial, in that observed changes in range are associated with greater-than-proportional changes in abundance. Although this relationship appears to be pervasive, and has important implications for the conservation of endangered species, the mechanism(s) underlying it remain poorly understood.
An ecological metacommunity is a set of interacting communities which are linked by the dispersal of multiple, potentially interacting species. The term is derived from the field of community ecology, which is primarily concerned with patterns of species distribution, abundance and interactions. Metacommunity ecology combines the importance of local factors and regional factors to explain patterns of species distributions that happen in different spatial scales.
Russell Scott Lande is an American evolutionary biologist and ecologist, and an International Chair Professor at Centre for Biodiversity Dynamics at the Norwegian University of Science and Technology (NTNU). He is a fellow of the Royal Society and a member of the United States National Academy of Sciences.
In ecology, extinction debt is the future extinction of species due to events in the past. The phrases dead clade walking and survival without recovery express the same idea.
The rescue effect is a phenomenon which was first described by Brown and Kodric-Brown, and is commonly used in metapopulation dynamics and many other disciplines in ecology. This populational process explains how the migration of individuals can increase the persistence of small isolated populations by helping to stabilize a metapopulation, thus reducing the chances of extinction. In other words, immigration can lead to the recolonization of previously extinct patches, promoting the long-term persistence of the network of populations.
Jordi Bascompte is a professor of ecology at the University of Zurich and the director of its specialized master's program on quantitative environmental sciences. He is best known for having brought the interactions of mutual benefit between plants and animals into community ecology, at the time largely dominated by predation and competition. His application of network theory to the study of mutualism has identified general laws that determine the way in which species interactions shape biodiversity.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.