Related Research Articles

Eureptilia is one of the two major subgroups of the clade Sauropsida, the other one being Parareptilia. Eureptilia includes Diapsida, as well as a number of primitive Permo-Carboniferous forms previously classified under Anapsida, in the old order "Cotylosauria".

Adelospondyli is an order of elongated, presumably aquatic, Carboniferous amphibians. They have a robust skull roofed with solid bone, and orbits located towards the front of the skull. The limbs were almost certainly absent, although some historical sources reported them to be present. Despite the likely absence of limbs, adelospondyls retained a large part of the bony shoulder girdle. Adelospondyls have been assigned to a variety of groups in the past. They have traditionally been seen as members of the subclass Lepospondyli, related to other unusual early tetrapods such as "microsaurs", "nectrideans", and aïstopods. Analyses such as Ruta & Coates (2007) have offered an alternate classification scheme, arguing that adelospondyls were actually far removed from other lepospondyls, instead being stem-tetrapod stegocephalians closely related to the family Colosteidae.

Westlothiana is a genus of reptile-like tetrapod that lived about 338 million years ago during the latest part of the Visean age of the Carboniferous. Members of the genus bore a superficial resemblance to modern-day lizards. The genus is known from a single species, Westlothiana lizziae. The type specimen was discovered in the East Kirkton Limestone at the East Kirkton Quarry, West Lothian, Scotland in 1984. This specimen was nicknamed "Lizzie the lizard" by fossil hunter Stan Wood, and this name was quickly adopted by other paleontologists and the press. When the specimen was formally named in 1990, it was given the specific name "lizziae" in homage to this nickname. However, despite its similar body shape, Westlothiana is not considered a true lizard. Westlothiana's anatomy contained a mixture of both "labyrinthodont" and reptilian features, and was originally regarded as the oldest known reptile or amniote. However, updated studies have shown that this identification is not entirely accurate. Instead of being one of the first amniotes, Westlothiana was rather a close relative of Amniota. As a result, most paleontologists since the original description place the genus within the group Reptiliomorpha, among other amniote relatives such as diadectomorphs and seymouriamorphs. Later analyses usually place the genus as the earliest diverging member of Lepospondyli, a collection of unusual tetrapods which may be close to amniotes or lissamphibians, or potentially both at the same time.
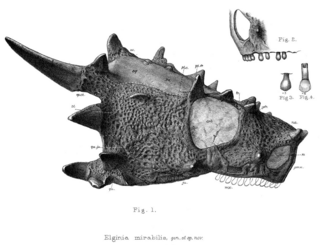
Elginia is an extinct genus of pareiasaurid known from the Late Permian of Scotland and China. It was named for the area around Elgin in Scotland, which has yielded many fossils referred to as the Elgin Reptiles.

Eosuchians are an extinct order of diapsid reptiles. Depending on which taxa are included the order may have ranged from the late Carboniferous to the Eocene but the consensus is that eosuchians are confined to the Permian and Triassic.

Diplocaulus is an extinct genus of lepospondyl amphibians which lived from the Late Carboniferous to the Late Permian of North America and Africa. Diplocaulus is by far the largest and best-known of the lepospondyls, characterized by a distinctive boomerang-shaped skull. Remains attributed to Diplocaulus have been found from the Late Permian of Morocco and represent the youngest-known occurrence of a lepospondyl.

Saharastega is an extinct genus of basal temnospondyl which lived during the Late Permian period, around 251 to 260 million years ago. Remains of Saharastega, discovered by paleontologist Christian A. Sidor at the Moradi Formation in Niger, were described briefly in 2005 and more comprehensively in 2006. The description is based on a skull lacking the lower jaws.

Rhinesuchidae is a family of tetrapods that lived primarily in the Permian period. They belonged to the broad group Temnospondyli, a successful and diverse collection of semiaquatic tetrapods which modern amphibians are probably descended from. Rhinesuchids can be differentiated from other temnospondyls by details of their skulls, most notably the interior structure of their otic notches at the back of the skull. They were among the earliest-diverging members of the Stereospondyli, a subgroup of temnospondyls with flat heads and aquatic habits. Although more advanced stereospondyls evolved to reach worldwide distribution in the Triassic period, rhinesuchids primarily lived in the high-latitude environments of Gondwana during the Guadalupian and Lopingian epochs of the Permian. The taxonomy of this family has been convoluted, with more than twenty species having been named in the past; a 2017 review recognized only eight of them to be valid. While several purported members of this group have been reported to have lived in the Triassic period, most are either dubious or do not belong to the group. However, at least one valid genus of rhinesuchid is known from the early Triassic, a small member known as Broomistega. The most recent formal definition of Rhinesuchidae, advocated by Mariscano et al. (2017) is that of a stem-based clade containing all taxa more closely related to Rhinesuchus whaitsi than to Lydekkerina huxleyi or Peltobatrachus pustulatus. A similar alternate definition is that Rhinesuchidae is a stem-based clade containing all taxa more closely related to Uranocentrodon senekalensis than to Lydekkerina huxleyi, Trematosaurus brauni, or Mastodonsaurus giganteus.

Acherontiscus is an extinct genus of stegocephalians that lived in the Early Carboniferous of Scotland. The type and only species is Acherontiscus caledoniae, named by paleontologist Robert Carroll in 1969. Members of this genus have an unusual combination of features which makes their placement within amphibian-grade tetrapods uncertain. They possess multi-bone vertebrae similar to those of embolomeres, but also a skull similar to lepospondyls. The only known specimen of Acherontiscus possessed an elongated body similar to that of a snake or eel. No limbs were preserved, and evidence for their presence in close relatives of Acherontiscus is dubious at best. Phylogenetic analyses created by Marcello Ruta and other paleontologists in the 2000s indicate that Acherontiscus is part of Adelospondyli, closely related to other snake-like animals such as Adelogyrinus and Dolichopareias. Adelospondyls are traditionally placed within the group Lepospondyli due to their fused vertebrae. Some analyses published since 2007 have argued that adelospondyls such as Acherontiscus may not actually be lepospondyls, instead being close relatives or members of the family Colosteidae. This would indicate that they evolved prior to the split between the tetrapod lineage that leads to reptiles (Reptiliomorpha) and the one that leads to modern amphibians (Batrachomorpha). Members of this genus were probably aquatic animals that were able to swim using snake-like movements.

Cryobatrachus is an extinct genus of temnospondyl amphibian from the Early Triassic of Antarctica. The type species is Cryobatrachus kitchingi. It is known from a partial skull and an imprint of the skull roof, both found in the Fremouw Formation of the Transantarctic Mountains at about 85° south latitude and described in 1974. Many small bone fragments have also been identified, although they cannot be attributed with certainty to C. kitchingi. Cryobatrachus has been classified in the family Lydekkerinidae, as it is similar in appearance to the genus Lydekkerina from South Africa. Because only a small number of features distinguish it from other lydekkerinids, Cryobatrachus kitchingi has more recently been considered a nomen dubium, meaning that its distinction from other better-known species may be unwarranted.
Rotaurisaurus is an extinct genus of amphibian-grade tetrapod from the family Lapillopsidae. This genus is known only from an incomplete crushed skull and associated left jaw, together given the designation UTGD 87795. The generic name, Rotaurisaurus, is a combination of Latin words translating to "circle-eared lizard". This references the shape of its otic notches, which acquire a circular form due to being partially enclosed by the tabular bones at the back of the skull. The specific name, contundo, references the specimen's poor level of preservation, as it is derived from the Latin word for "squashed".

Scincosaurus is an extinct genus of nectridean lepospondyl within the family Scincosauridae.

Subcyclotosaurus is an extinct genus of carnivorous prehistoric mastodonsaurid temnospondyl amphibian. It is known from a single partial skull from the Hawkesbury Sandstone formation in Australia.

Biseridens is an extinct genus of anomodont therapsid, and one of the most basal anomodont genera known. Originally known from a partial skull misidentified as an 'eotitanosuchian' in 1997, another well-preserved skull was found in the Xidagou Formation, an outcropping in the Qilian Mountains of Gansu, China, in 2009 that clarified its relationships to anomodonts, such as the dicynodonts.
Deltaherpeton is an extinct genus of colosteid from middle Mississippian deposits of Delta, Iowa, United States. It was first named by John R. Bolt and R. Eric Lombard in 2010 and the type species is Deltaherpeton hiemstrae.
Procyclotosaurus is an extinct genus of stenotosaurid capitosaurian temnospondyl. The type species is P. stantonensis. In 1904, English paleontologist Arthur Smith Woodward described it as a species of Capitosaurus, C. stantonensis, based on a partial skull known as R 3174. In 1958, the species was assigned to the new genus. It is known from the Lower Keuper, a European stratigraphic unit that was deposited during the late Middle Triassic. Fossils have been found from Staffordshire, England.

Postparietals are cranial bones present in fish and many tetrapods. Although initially a pair of bones, many lineages possess postparietals which were fused into a single bone. The postparietals were dermal bones situated along the midline of the skull, behind the parietal bones. They formed part of the rear edge of the skull roof, and the lateral edge of each postparietal often contacts the tabular and supratemporal bones. In fish, the postparietals are elongated, typically the largest components of the skull roof. Tetrapods possessed shorter postparietals, which were reduced further and shifted towards the braincase in amniotes. At several points in synapsid evolution, the postparietals fused to each other and the tabulars during embryological development. This fusion produces the interparietal bone, which is inherited by mammals. Postparietals are common in extinct amphibians and early reptiles. However, most living amphibians and living reptiles lack postparietal bones, with a few exceptions.
Ossirarus is an extinct genus of four-limbed stem-tetrapod from the Mississippian (mid-Tournaisian) of Scotland. It contains a single species, Ossirarus kierani, based on disarticulated skull and postcranial bones from the Ballagan Formation at Burnmouth. It would have had a large, pointed tabular horn and several minor traits shared with Devonian stem-tetrapods. Ossirarus was described in a 2016 study which was devised to fill in the tetrapod and stem-tetrapod faunas of Romer's gap, an interval of the early Carboniferous with few vertebrate fossils. It was one of five new genera named in this study, along with Aytonerpeton, Diploradus, Koilops, and Perittodus.
The supratemporal bone is a paired cranial bone present in many tetrapods and tetrapodomorph fish. It is part of the temporal region, usually lying medial (inwards) relative to the squamosal and lateral (outwards) relative to the parietal and/or postparietal. It may also contact the postorbital or intertemporal, or tabular, when those bones are present.
References
- ↑ taboo word tabulate. "📌 tabular bone". Enacademic.com. Retrieved 2019-11-21.
| | This fish-related article is a stub. You can help Wikipedia by expanding it. |
| | This amphibian-related article is a stub. You can help Wikipedia by expanding it. |
| | This vertebrate anatomy–related article is a stub. You can help Wikipedia by expanding it. |