Catechol 1,2- dioxygenase (EC1.13.11.1, 1,2-CTD, catechol-oxygen 1,2-oxidoreductase, 1,2-pyrocatechase, catechase, catechol 1,2-oxygenase, catechol dioxygenase, pyrocatechase, pyrocatechol 1,2-dioxygenase, CD I, CD II)[1][2][3][4] is an enzyme that catalyzes the oxidative ring cleavage of catechol to form cis,cis-muconic acid:[5]
Figure 1. The overall reaction of catechol 1,2-dioxygenase. Using a non-heme iron(III) complex, 1,2-CTD is able to oxidatively cleave catechol into cis,cis-muconic acid.
More specifically, 1,2-CTD is an intradiol dioxygenase, a family of catechol dioxygenases that cleaves the bond between the phenolic hydroxyl groups of catechol using an Fe3+ cofactor.
Two families of dioxygenases were discovered by Osamu Hayaishi and Kizo Hashimoto in 1950: catechol 1,2-dioxygenase and catechol 2,3-dioxygenase (2,3-CTD).[22] The two enzymes were identified to be a part of two separate catechol dioxygenase families: 1,2-CTD was classified as an intradiol dioxygenase while 2,3-CTD was classified as an extradiol dioxygenase. The two enzymes can be distinguished based on their reaction products and cofactors. 1,2-CTD uses Fe3+ as a cofactor to cleave the carbon-carbon bond between the phenolic hydroxyl groups of catechol, thus yielding muconic acid as its product. In contrast, 2,3-CTD utilizes Fe2+ as a cofactor to cleave the carbon-carbon bond adjacent to the phenolic hydroxyl groups of catechol, thus yielding 2-hydroxymuconaldehye as its product.[23]
Enzyme Structure
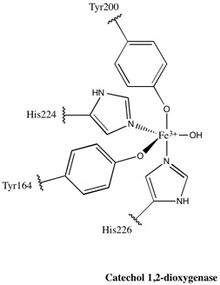
Figure 2. The active site of 1,2-CTD. This non-heme Fe complex is axially ligated by Tyr200 and His226 and equatorially ligated by Tyr164, His224, and a solvent water molecule, giving the active site an overall trigonal bipyramidal geometry
Almost all members of the 1,2-CTD family are homodimers; the 1,2-CTD enzyme produced by Pseudomonas arvilla is the exception to this rule, containing two highly homologous subunits that can form either a homo- or hetero- dimer.[24] The enzyme resembles a boomerang in shape, and can therefore be clearly divided into three domains: two catalytic domains residing at each end of the “boomerang” and a linker domain at the center.
Each catalytic domain is composed of two stacked, mixed topology β sheets and several random coils. These sheets and coils subsequently encompass the active site: a non-heme iron(III) complex. Without heme, iron must be ligated to four amino acid residues (Tyr200, His226, Tyr164, His224) to maintain is catalytically active conformation. With Tyr200 and His226 acting as the axial ligands and Tyr164, His224, and a solvent water molecule acting as equatorial ligands, the Fe3+ complex displays trigonal bipyramidal geometry. Since the active sites of each catalytic domain are separated by a distance of 40 Å, they are not believed to allosterically effect one another.
In contrast, the linker domain is composed of α helices supplied by the two catalytic domains: each domain contributes five helices from their N termini and one from a helix that spans both the catalytic domain and the linker domain. At the center of the linker domain resides an 8 by 35 Å hydrophobic tunnel with two phospholipids bound at each end. The head of each phospholipid points outward towards solution while the tails are embedded within the enzyme. The function of this hydrophobic tunnel is unknown, though two hypotheses have been postulated concerning its utility. The first is that the binding of the terminal phospholipids alters the conformation of the active sites, implying that the tunnel acts as an effector, only allowing the enzyme to be active in certain areas of the cell. The second hypothesis postulates that the tunnel regulates lipid membrane rigidity through its degradation of phenolic hydrocarbons and ability to bind to other lipids. Studies have shown that phenolic hydrocarbons affect the functional and structural properties of cell membranes.[25] 1,2-CTD degrades phenolic hydrocarbons key to the synthesis lipid membranes. Therefore, 1,2-CTD may bind to the cell lipid membrane via its terminal phospholipids and thus have greater access to the phenolic hydrocarbons vital in lipid membrane structure.[26]
Enzyme Mechanism
Figure 3. The mechanism by which 1,2-CTD utilizes a non-heme Fe complex to oxidatively cleave catechol to form cis,cis-muconic acid.
The catalytic mechanism of catechol 1,2-dioxygenase was elucidated using a combination of O18 labeling experiments and crystallography.[27][28][29] Upon entering the active site, the hydroxyl group on the fourth carbon (C4) of catechol binds to Fe3+; this binding is facilitated by the hydroxide ligand, which deprotonates the C4 hydroxyl group. The second catechol hydroxyl group on carbon 3 (C3) is coordinated to Fe3+ after its deprotonation by the Tyr200 ligand. In the process of deprotonating the C3 hydroxyl group, Tyr200 dissociates from the catalytic complex.
Oxygen is bonded to the substrate through a series of trans influences and stabilizing hydrogen bonding between the substrate and other active site amino acid residues. His226 accepts electron density from the substrate, consequently decreasing the bond between Fe3+ and the C4 hydroxyl. At the same time the bond between the C3 hydroxyl and Fe3+ is increased due to the electron withdrawing effects of Tyr164. These distortions, coupled with the hydrogen bonding between Arg221 and the C3 hydroxyl, induces the C3 hydroxyl group to ketonize and increases the carbanion character of C4. The newly formed C4 carbanion attacks O2, thus binding it to the substrate.
Another trans influence follows the aforementioned step, leading to the cleavage of O2 and the subsequent insertion of one of the oxygen molecules between C3 and C4. Rapid hydrolysis follows this reaction, yielding a primed active site and product.
Related Research Articles
Catechol, also known as pyrocatechol or 1,2-dihydroxybenzene, is an organic compound with the molecular formula C6H4(OH)2. It is the ortho isomer of the three isomeric benzenediols. This colorless compound occurs naturally in trace amounts. It was first discovered by destructive distillation of the plant extract catechin. About 20,000 tonnes of catechol are now synthetically produced annually as a commodity organic chemical, mainly as a precursor to pesticides, flavors, and fragrances. Small amounts of catechol occur in fruits and vegetables.
Aromatic-ring-hydroxylating dioxygenases (ARHD) incorporate two atoms of dioxygen (O2) into their substrates in the dihydroxylation reaction. The product is (substituted) cis-1,2-dihydroxycyclohexadiene, which is subsequently converted to (substituted) benzene glycol by a cis-diol dehydrogenase.
Lipoxygenases (LOX) are a family of (non-heme) iron-containing enzymes, more specifically oxidative enzymes, most of which catalyze the dioxygenation of polyunsaturated fatty acids in lipids containing a cis,cis-1,4-pentadiene into cell signaling agents that serve diverse roles as autocrine signals that regulate the function of their parent cells, paracrine signals that regulate the function of nearby cells, and endocrine signals that regulate the function of distant cells.
Phosphoinositide phospholipase C is a family of eukaryotic intracellular enzymes that play an important role in signal transduction processes. These enzymes belong to a larger superfamily of Phospholipase C. Other families of phospholipase C enzymes have been identified in bacteria and trypanosomes. Phospholipases C are phosphodiesterases.
An oxygenase is any enzyme that oxidizes a substrate by transferring the oxygen from molecular oxygen O2 (as in air) to it. The oxygenases form a class of oxidoreductases; their EC number is EC 1.13 or EC 1.14.
Methane monooxygenase (MMO) is an enzyme capable of oxidizing the C-H bond in methane as well as other alkanes. Methane monooxygenase belongs to the class of oxidoreductase enzymes.
Catechol dioxygenases are metalloprotein enzymes that carry out the oxidative cleavage of catechols. This class of enzymes incorporate dioxygen into the substrate. Catechol dioxygenases belong to the class of oxidoreductases and have several different substrate specificities, including catechol 1,2-dioxygenase, catechol 2,3-dioxygenase, and protocatechuate 3,4-dioxygenase. The active site of catechol dioxygenases most frequently contains iron, but manganese-containing forms are also known.
Catechol 2,3-dioxygenase (EC 1.13.11.2, 2,3-pyrocatechase, catechol 2,3-oxygenase, catechol oxygenase, metapyrocatechase, pyrocatechol 2,3-dioxygenase) is an enzyme with systematic name catechol:oxygen 2,3-oxidoreductase (decyclizing). This enzyme catalyses the following chemical reaction
In enzymology, a 2-chlorobenzoate 1,2-dioxygenase (EC 1.14.12.13) is an enzyme that catalyzes the chemical reaction
4-hydroxyphenylacetate 3-monooxygenase (EC 1.14.14.9) is an enzyme that catalyzes the chemical reaction
In enzymology, an anthranilate 1,2-dioxygenase (deaminating, decarboxylating) (EC 1.14.12.1) is an enzyme that catalyzes the chemical reaction
In enzymology, a benzoate 1,2-dioxygenase (EC 1.14.12.10) is an enzyme that catalyzes the chemical reaction
In enzymology, a biphenyl 2,3-dioxygenase (EC 1.14.12.18) is an enzyme that catalyzes the chemical reaction
Gamma-butyrobetaine dioxygenase is an enzyme that in humans is encoded by the BBOX1 gene. Gamma-butyrobetaine dioxygenase catalyses the formation of L-carnitine from gamma-butyrobetaine, the last step in the L-carnitine biosynthesis pathway. Carnitine is essential for the transport of activated fatty acids across the mitochondrial membrane during mitochondrial beta oxidation. In humans, gamma-butyrobetaine dioxygenase can be found in the kidney (high), liver (moderate), and brain. BBOX1 has recently been identified as a potential cancer gene based on a large-scale microarray data analysis.
In enzymology, a naphthalene 1,2-dioxygenase (EC 1.14.12.12) is an enzyme that catalyzes the chemical reaction
Chloridazon-catechol dioxygenase (EC 1.13.11.36) is an enzyme that catalyzes the chemical reaction
In enzymology, a gentisate 1,2-dioxygenase (EC 1.13.11.4) is an enzyme that catalyzes the chemical reaction
In enzymology, a protocatechuate 3,4-dioxygenase (EC 1.13.11.3) is an enzyme that catalyzes the chemical reaction
Dioxygenases are oxidoreductase enzymes. Aerobic life, from simple single-celled bacteria species to complex eukaryotic organisms, has evolved to depend on the oxidizing power of dioxygen in various metabolic pathways. From energetic adenosine triphosphate (ATP) generation to xenobiotic degradation, the use of dioxygen as a biological oxidant is widespread and varied in the exact mechanism of its use. Enzymes employ many different schemes to use dioxygen, and this largely depends on the substrate and reaction at hand.
3-Methylcatechol is an organic compound with the formula CH3C6H3(OH)2 A white solid, it is one of the isomers of methylbenzenediol. Being structurally related to lignans, it is contributes to the aerosol generate by combustion of wood.
References
↑ Hayaishi O, Lardy H, Myrbäck K (1963). "Direct oxygenation by O2, oxygenases". In Boyer PD (ed.). The Enzymes. Vol.8 (2nded.). New York: Academic Press. pp.353–371.
↑ Hayaishi O, Katagiri M, Rothberg S (December 1957). "Studies on oxygenases; pyrocatechase". The Journal of Biological Chemistry. 229 (2): 905–20. PMID13502352.
↑ Sistrom WR, Stanier RY (October 1954). "The mechanism of formation of beta-ketoadipic acid by bacteria". The Journal of Biological Chemistry. 210 (2): 821–36. PMID13211620.
↑ Itoh, M (1981). "Characteristics of a new catechol-1,2-oxygenase from Trichosporon cutaneum WY2-2". Agric. Biol. Chem. 45 (1): 2787–2796.
↑ Aoki K, Konohana T, Shinke R, Nishira H (1984). "Classification of catechol 1,2-dioxygenase family: sequence analysis of a gene for the catechol 1,2-dioxygenase showing high specificity for methylcatechols from Gram+ aniline-assimilating Rhodococcus erythropolis AN-13". Agric. Biol. Chem. 48 (1): 2087–2095. doi:10.1016/S0378-1119(96)00629-4. PMID9034312.
↑ Aoki K, Konohana T, Shinke R, Nishira H (1984). "Two catechol 1,2-dioxygenases from aniline-assimilating bacterium, Frateuria species ANA-18". Agric. Biol. Chem. 48 (1): 2097–2104.
↑ Chen Y, Glenn A, Dilworth M (1985). "Aromatic metabolism in Rhizobium trifolii-catechol 1,2-dioxygenase". Arch. Microbiol. 141 (1): 225–228. doi:10.1007/BF00408063.
↑ Pascal RA, Huang DS (July 1986). "Reactions of 3-ethylcatechol and 3-(methylthio)catechol with catechol dioxygenases". Archives of Biochemistry and Biophysics. 248 (1): 130–7. doi:10.1016/0003-9861(86)90409-1. PMID3015028.
↑ Krug M, Straube G (1986). "Degradation of phenolic compounds by the yeast Candida tropicalis HP 15. II. Some properties of the first two enzymes of the degradation pathway". Journal of Basic Microbiology. 26 (5): 271–81. doi:10.1002/jobm.3620260505. PMID3783431.
↑ Smith M, Ratledge C, Crook S (1990). "Properties of cyanogen bromide-activated, Agarose-immobilized catechol 1,2-dioxygenase from freeze-dried extracts of Nocardia sp. NCIB 10503". Enzyme Microb. Technol. 12 (12): 945–949. doi:10.1016/0141-0229(90)90114-6.
↑ Que L, Ho RY (November 1996). "Dioxygen Activation by Enzymes with Mononuclear Non-Heme Iron Active Sites". Chemical Reviews. 96 (7): 2607–2624. doi:10.1021/cr960039f. PMID11848838.
↑ Hayaishi O, Hashimoto K (1950). "Pyrocatecase a New Enzyme Catalizing Oxidative Breakdown of Pyrocatechin". J. Biochem. 37 (3): 371–374. doi:10.1093/oxfordjournals.jbchem.a126205.
↑ Bugg, Timothy (2003). "Dioxygenase enzymes: catalytic mechanisms and chemical models". Tetrahedron. 59 (36): 7075–7101. doi:10.1016/S0040-4020(03)00944-X.
↑ Nakai C, Kagamiyama H, Saeki Y, Nozaki M (June 1979). "Nonidentical subunits of pyrocatechase from Pseudomonas arvilla C-1". Archives of Biochemistry and Biophysics. 195 (1): 12–22. doi:10.1016/0003-9861(79)90322-9. hdl:2433/222081. PMID475379.
↑ Hayaishi O, Hashimoto K (1950). "Pyrocatecase a New Enzyme Catalizing Oxidative Breakdown of Pyrocatechin". J. Biochem. 37 (3): 371–374. doi:10.1093/oxfordjournals.jbchem.a126205.
↑ Orville AM, Lipscomb JD, Ohlendorf DH (August 1997). "Crystal structures of substrate and substrate analog complexes of protocatechuate 3,4-dioxygenase: endogenous Fe3+ ligand displacement in response to substrate binding". Biochemistry. 36 (33): 10052–66. doi:10.1021/bi970469f. PMID9254600.
↑ Lipscomb J, Orville A (1992). Metal Ions in Biological Systems. Vol.28. Marcel Dekker, Inc. pp.243–298.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.