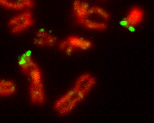
Region of sexual chromosomes exhibiting an autosomal inheritance pattern
Detail of a human metaphase spread. A region in the pseudoautosomal region of the short arms of the X chromosome (left) and the Y chromosome (top right) was detected by fluorescent in situ hybridization (green). Chromosomes counterstained in red.
The pseudoautosomal regions get their name because any genes within them (so far at least 29 have been found for humans)[3] are inherited just like any autosomal genes. In humans, these regions are referred to as PAR1 and PAR2.[4] PAR1 comprises 2.6 Mbp of the short-arm tips of both X and Y chromosomes in humans and great apes (X and Y are 154 Mbp and 62 Mbp in total). PAR2 is at the tips of the long arms, spanning 320 kbp.[5] The monotremes, including the platypus and echidna, have a multiple sex chromosome system, and consequently have 8 pseudoautosomal regions.[6]
Location
Pseudoautosomal regions are at both termini of the sex chromosomes.
Normal male therian mammals have two copies of these genes: one in the pseudoautosomal region of their Y chromosome, the other in the corresponding portion of their X chromosome. Normal females also possess two copies of pseudoautosomal genes, as each of their two X chromosomes contains a pseudoautosomal region. Crossing over between the X and Y chromosomes is normally restricted to the pseudoautosomal regions; thus, pseudoautosomal genes exhibit an autosomal, rather than sex-linked, pattern of inheritance. So, females can inherit an allele originally present on the Y chromosome of their father.
The function of these pseudoautosomal regions is that they allow the X and Y chromosomes to pair and properly segregate during meiosis in males.[9]
Genes
PAR1 contains 16 genes, with PLCXD1 as the furthermost PAR1 gene at the distal telomeric end and XG at the boundary of PAR1 at the centromeric end. PAR2 contains 3 genes, with SPRY3 at the centromeric boundary and IL9R at the distal telomeric end.
Pseudoautosomal genes are found in two different locations: PAR1 and PAR2. These are believed to have evolved independently.[11]
Pairing (synapsis) of the X and Y chromosomes and crossing over (recombination) between their pseudoautosomal regions appear to be necessary for the normal progression of male meiosis.[16] Thus, those cells in which X-Y recombination does not occur will fail to complete meiosis. Structural and/or genetic dissimilarity (due to hybridization or mutation) between the pseudoautosomal regions of the X and Y chromosomes can disrupt pairing and recombination, and consequently cause male infertility.
The SHOX gene in the PAR1 region is the gene most commonly associated with and well understood with regards to disorders in humans,[17] but all pseudoautosomal genes escape X-inactivation and are therefore candidates for having gene dosage effects in sex chromosome aneuploidy conditions (45,X, 47,XXX, 47,XXY, 47,XYY, etc.).
↑ "WASH6P". HUGO Gene Nomenclature Committee. Retrieved 2019-08-30.
↑ Eichner, E.M. (February 1991). "The mouse Y* chromosome involves a complex rearrangement including interstitial positioning of the Y-pseudoautosomal region". Cytogenetics and Cell Genetics. 57 (4): 221–230. doi:10.1159/000133152. PMID1743079.
↑ Blaschke RJ, Rappold G (June 2006). "The pseudoautosomal regions, SHOX and disease". Curr. Opin. Genet. Dev. 16 (3): 233–9. doi:10.1016/j.gde.2006.04.004. PMID16650979.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.