
Queueing theory is the mathematical study of waiting lines, or queues. A queueing model is constructed so that queue lengths and waiting time can be predicted. Queueing theory is generally considered a branch of operations research because the results are often used when making business decisions about the resources needed to provide a service.
The laser diode rate equations model the electrical and optical performance of a laser diode. This system of ordinary differential equations relates the number or density of photons and charge carriers (electrons) in the device to the injection current and to device and material parameters such as carrier lifetime, photon lifetime, and the optical gain.
The birth–death process is a special case of continuous-time Markov process where the state transitions are of only two types: "births", which increase the state variable by one and "deaths", which decrease the state by one. The model's name comes from a common application, the use of such models to represent the current size of a population where the transitions are literal births and deaths. Birth–death processes have many applications in demography, queueing theory, performance engineering, epidemiology, biology and other areas. They may be used, for example, to study the evolution of bacteria, the number of people with a disease within a population, or the number of customers in line at the supermarket.
In mathematics and probability theory, a gamma process, also known as (Moran-)Gamma subordinator, is a random process with independent gamma distributed increments. Often written as , it is a pure-jump increasing Lévy process with intensity measure for positive . Thus jumps whose size lies in the interval occur as a Poisson process with intensity The parameter controls the rate of jump arrivals and the scaling parameter inversely controls the jump size. It is assumed that the process starts from a value 0 at t = 0.
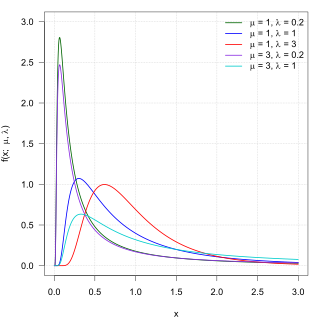
In probability theory, the inverse Gaussian distribution is a two-parameter family of continuous probability distributions with support on (0,∞).
In probability theory relating to stochastic processes, a Feller process is a particular kind of Markov process.
The maximal ergodic theorem is a theorem in ergodic theory, a discipline within mathematics.
The linear attenuation coefficient, attenuation coefficient, or narrow-beam attenuation coefficient characterizes how easily a volume of material can be penetrated by a beam of light, sound, particles, or other energy or matter. A coefficient value that is large represents a beam becoming 'attenuated' as it passes through a given medium, while a small value represents that the medium had little effect on loss. The SI unit of attenuation coefficient is the reciprocal metre (m−1). Extinction coefficient is another term for this quantity, often used in meteorology and climatology. Most commonly, the quantity measures the exponential decay of intensity, that is, the value of downward e-folding distance of the original intensity as the energy of the intensity passes through a unit thickness of material, so that an attenuation coefficient of 1 m−1 means that after passing through 1 metre, the radiation will be reduced by a factor of e, and for material with a coefficient of 2 m−1, it will be reduced twice by e, or e2. Other measures may use a different factor than e, such as the decadic attenuation coefficient below. The broad-beam attenuation coefficient counts forward-scattered radiation as transmitted rather than attenuated, and is more applicable to radiation shielding.
In queueing theory, a discipline within the mathematical theory of probability, the Pollaczek–Khinchine formula states a relationship between the queue length and service time distribution Laplace transforms for an M/G/1 queue. The term is also used to refer to the relationships between the mean queue length and mean waiting/service time in such a model.

In queueing theory, a discipline within the mathematical theory of probability, an M/M/1 queue represents the queue length in a system having a single server, where arrivals are determined by a Poisson process and job service times have an exponential distribution. The model name is written in Kendall's notation. The model is the most elementary of queueing models and an attractive object of study as closed-form expressions can be obtained for many metrics of interest in this model. An extension of this model with more than one server is the M/M/c queue.
In queueing theory, a discipline within the mathematical theory of probability, the M/M/c queue is a multi-server queueing model. In Kendall's notation it describes a system where arrivals form a single queue and are governed by a Poisson process, there are c servers, and job service times are exponentially distributed. It is a generalisation of the M/M/1 queue which considers only a single server. The model with infinitely many servers is the M/M/∞ queue.
In probability theory, a Laplace functional refers to one of two possible mathematical functions of functions or, more precisely, functionals that serve as mathematical tools for studying either point processes or concentration of measure properties of metric spaces. One type of Laplace functional, also known as a characteristic functional is defined in relation to a point process, which can be interpreted as random counting measures, and has applications in characterizing and deriving results on point processes. Its definition is analogous to a characteristic function for a random variable.
In queueing theory, a discipline within the mathematical theory of probability, the backpressure routing algorithm is a method for directing traffic around a queueing network that achieves maximum network throughput, which is established using concepts of Lyapunov drift. Backpressure routing considers the situation where each job can visit multiple service nodes in the network. It is an extension of max-weight scheduling where each job visits only a single service node.
In queueing theory, a discipline within the mathematical theory of probability, a fluid queue is a mathematical model used to describe the fluid level in a reservoir subject to randomly determined periods of filling and emptying. The term dam theory was used in earlier literature for these models. The model has been used to approximate discrete models, model the spread of wildfires, in ruin theory and to model high speed data networks. The model applies the leaky bucket algorithm to a stochastic source.
In queueing theory, a discipline within the mathematical theory of probability, the M/M/∞ queue is a multi-server queueing model where every arrival experiences immediate service and does not wait. In Kendall's notation it describes a system where arrivals are governed by a Poisson process, there are infinitely many servers, so jobs do not need to wait for a server. Each job has an exponentially distributed service time. It is a limit of the M/M/c queue model where the number of servers c becomes very large.
In queueing theory, a discipline within the mathematical theory of probability, an M/D/1 queue represents the queue length in a system having a single server, where arrivals are determined by a Poisson process and job service times are fixed (deterministic). The model name is written in Kendall's notation. Agner Krarup Erlang first published on this model in 1909, starting the subject of queueing theory. An extension of this model with more than one server is the M/D/c queue.
In probability theory, a transition rate matrix is an array of numbers describing the instantaneous rate at which a continuous time Markov chain transitions between states.
Biogeography-based optimization (BBO) is an evolutionary algorithm (EA) that optimizes a function by stochastically and iteratively improving candidate solutions with regard to a given measure of quality, or fitness function. BBO belongs to the class of metaheuristics since it includes many variations, and since it does not make any assumptions about the problem and can therefore be applied to a wide class of problems.
Diversification rates are the rates at which new species form and living species go extinct. Diversification rates can be estimated from fossils, data on the species diversity of clades and their ages, or phylogenetic trees. Diversification rates are typically reported on a per-lineage basis, and refer to the diversification dynamics expected under a birth–death process.
A mixed Poisson distribution is a univariate discrete probability distribution in stochastics. It results from assuming that a random variable is Poisson distributed, where the rate parameter itself is considered as a random variable. Hence it is a special case of a compound probability distribution. Mixed Poisson distributions can be found in actuarial mathematics as a general approach for the distribution of the number of claims and is also examined as an epidemiological model. It should not be confused with compound Poisson distribution or compound Poisson process.










