
Parasitism is a close relationship between species, where one organism, the parasite, lives on or inside another organism, the host, causing it some harm, and is adapted structurally to this way of life. The entomologist E. O. Wilson characterised parasites as "predators that eat prey in units of less than one". Parasites include single-celled protozoans such as the agents of malaria, sleeping sickness, and amoebic dysentery; animals such as hookworms, lice, mosquitoes, and vampire bats; fungi such as honey fungus and the agents of ringworm; and plants such as mistletoe, dodder, and the broomrapes.
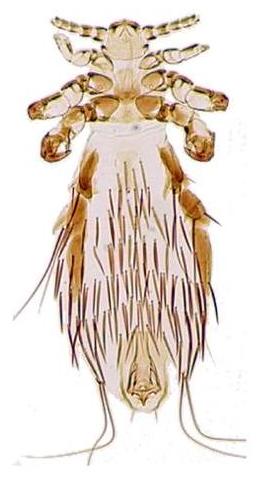
Louse is the common name for any member of the clade Phthiraptera, which contains nearly 5,000 species of wingless parasitic insects. Phthiraptera has variously been recognized as an order, infraorder, or a parvorder, as a result of developments in phylogenetic research.

In biology, coevolution occurs when two or more species reciprocally affect each other's evolution through the process of natural selection. The term sometimes is used for two traits in the same species affecting each other's evolution, as well as gene-culture coevolution.
Coextinction and cothreatened refer to the phenomenon of the loss or decline of a host species resulting in the loss or endangerment of another species that depends on it, potentially leading to cascading effects across trophic levels. The term was originated by the authors Stork and Lyal (1993) and was originally used to explain the extinction of parasitic insects following the loss of their specific hosts. The term is now used to describe the loss of any interacting species, including competition with their counterpart, and specialist herbivores with their food source. Coextinction is especially common when a keystone species goes extinct.
The Prodoxidae are a family of moths, generally small in size and nondescript in appearance. They include species of moderate pest status, such as the currant shoot borer, and others of considerable ecological and evolutionary interest, such as various species of "yucca moths".

Ant–fungus mutualism is a symbiosis seen between certain ant and fungal species, in which ants actively cultivate fungus much like humans farm crops as a food source. There is only evidence of two instances in which this form of agriculture evolved in ants resulting in a dependence on fungi for food. These instances were the attine ants and some ants that are part of the Megalomyrmex genus. In some species, the ants and fungi are dependent on each other for survival. This type of codependency is prevalent among herbivores who rely on plant material for nutrition. The fungus’ ability to convert the plant material into a food source accessible to their host makes them the ideal partner. The leafcutter ant is a well-known example of this symbiosis. Leafcutter ants species can be found in southern South America up to the United States. However, ants are not the only ground-dwelling arthropods which have developed symbioses with fungi. A similar mutualism with fungi is also noted in termites within the subfamily Macrotermitinae which are widely distributed throughout the Old World tropics with the highest diversity in Africa.

Roderic Dugald Morton Page is a New Zealand-born evolutionary biologist at the University of Glasgow, Scotland, and the author of several books. As of 2015 he is professor at the University of Glasgow and was editor of the journal Systematic Biology until the end of 2007. His main interests are in phylogenetics, evolutionary biology and bioinformatics.
The Red Queen's hypothesis is a hypothesis in evolutionary biology proposed in 1973, that species must constantly adapt, evolve, and proliferate in order to survive while pitted against ever-evolving opposing species. The hypothesis was intended to explain the constant (age-independent) extinction probability as observed in the paleontological record caused by co-evolution between competing species; however, it has also been suggested that the Red Queen hypothesis explains the advantage of sexual reproduction at the level of individuals, and the positive correlation between speciation and extinction rates in most higher taxa.
Avian malaria is a parasitic disease of birds, caused by parasite species belonging to the genera Plasmodium and Hemoproteus. The disease is transmitted by a dipteran vector including mosquitoes in the case of Plasmodium parasites and biting midges for Hemoproteus. The range of symptoms and effects of the parasite on its bird hosts is very wide, from asymptomatic cases to drastic population declines due to the disease, as is the case of the Hawaiian honeycreepers. The diversity of parasites is large, as it is estimated that there are approximately as many parasites as there are species of hosts. As research on human malaria parasites became difficult, Dr. Ross studied avian malaria parasites. Co-speciation and host switching events have contributed to the broad range of hosts that these parasites can infect, causing avian malaria to be a widespread global disease, found everywhere except Antarctica.

Entorrhizomycetes is the sole class in the phylum Entorrhizomycota, within the Fungi subkingdom Dikarya along with Basidiomycota and Ascomycota. It contains three genera and is a small group of teliosporic root parasites that form galls on plants in the Juncaceae (rush) and Cyperaceae (sedge) families. Prior to 2015 this phylum was placed under the subdivision Ustilaginomycotina. A 2015 study did a "comprehensive five-gene analyses" of Entorrhiza and concluded that the former class Entorrhizomycetes is possibly either a close sister group to the rest of Dikarya or Basidiomycota.
Host–parasite coevolution is a special case of coevolution, where a host and a parasite continually adapt to each other. This can create an evolutionary arms race between them. A more benign possibility is of an evolutionary trade-off between transmission and virulence in the parasite, as if it kills its host too quickly, the parasite will not be able to reproduce either. Another theory, the Red Queen hypothesis, proposes that since both host and parasite have to keep on evolving to keep up with each other, and since sexual reproduction continually creates new combinations of genes, parasitism favours sexual reproduction in the host.

Ecological fitting is "the process whereby organisms colonize and persist in novel environments, use novel resources or form novel associations with other species as a result of the suites of traits that they carry at the time they encounter the novel condition". It can be understood as a situation in which a species' interactions with its biotic and abiotic environment seem to indicate a history of coevolution, when in actuality the relevant traits evolved in response to a different set of biotic and abiotic conditions.
Incomplete lineage sorting (ILS) (also referred to as hemiplasy, deep coalescence, retention of ancestral polymorphism, or trans-species polymorphism) is a phenomena in evolutionary biology and population genetics that results in discordance between species and gene trees. By contrast, complete lineage sorting results in concordant species and gene trees. ILS occurs in the context of a gene in an ancestral species which exists in multiple alleles. If a speciation event occurs in this situation, either complete lineage sorting will occur, and both daughter species will inherit all alleles of the gene in question, or incomplete lineage sorting will occur, when one or both daughter species inherits a subset of alleles present in the parental species. For example, if two alleles of a gene are present and a speciation event occurs, one of the two daughter species might inherit both alleles, but the second daughter species only inherits one of the two alleles. In this case, incomplete lineage sorting has occurred.

Eichler's rule is one of several coevolutionary rules which states that parasites tend to be highly specific to their hosts, and thus it seems reasonable to expect a positive co-variation between the taxonomic richness of hosts and that of their parasites.

Escape and radiate coevolution is a hypothesis proposing that a coevolutionary 'arms-race' between primary producers and their consumers contributes to the diversification of species by accelerating speciation rates. The hypothesized process involves the evolution of novel defenses in the host, allowing it to "escape" and then "radiate" into differing species.

Ecological speciation is a form of speciation arising from reproductive isolation that occurs due to an ecological factor that reduces or eliminates gene flow between two populations of a species. Ecological factors can include changes in the environmental conditions in which a species experiences, such as behavioral changes involving predation, predator avoidance, pollinator attraction, and foraging; as well as changes in mate choice due to sexual selection or communication systems. Ecologically-driven reproductive isolation under divergent natural selection leads to the formation of new species. This has been documented in many cases in nature and has been a major focus of research on speciation for the past few decades.

A biological rule or biological law is a generalized law, principle, or rule of thumb formulated to describe patterns observed in living organisms. Biological rules and laws are often developed as succinct, broadly applicable ways to explain complex phenomena or salient observations about the ecology and biogeographical distributions of plant and animal species around the world, though they have been proposed for or extended to all types of organisms. Many of these regularities of ecology and biogeography are named after the biologists who first described them.

The genus Ficus is composed of 800 species of vines, shrubs, and trees, defined by their syconiums, the fruit-like vessels that either hold female flowers or pollen on the inside. In addition to being cultivated by humans for thousands of years, Ficus is also known for their reproductive mutualism with the fig wasp.

Harrison's rule is an observation in evolutionary biology by Launcelot Harrison which states that in comparisons across closely related species, host and parasite body sizes tend to covary positively.

In phylogenetics, reconciliation is an approach to connect the history of two or more coevolving biological entities. The general idea of reconciliation is that a phylogenetic tree representing the evolution of an entity can be drawn within another phylogenetic tree representing an encompassing entity to reveal their interdependence and the evolutionary events that have marked their shared history. The development of reconciliation approaches started in the 1980s, mainly to depict the coevolution of a gene and a genome, and of a host and a symbiont, which can be mutualist, commensalist or parasitic. It has also been used for example to detect horizontal gene transfer, or understand the dynamics of genome evolution.


