The Enzyme Commission number is a numerical classification scheme for enzymes, based on the chemical reactions they catalyze. As a system of enzyme nomenclature, every EC number is associated with a recommended name for the respective enzyme.
Catalysis is the process of increasing the rate of a chemical reaction by adding a substance known as a catalyst, which is not consumed in the catalyzed reaction and can continue to act repeatedly. Because of this, only very small amounts of catalyst are required to alter the reaction rate in principle.
A chemical reaction is a process that leads to the chemical transformation of one set of chemical substances to another. Classically, chemical reactions encompass changes that only involve the positions of electrons in the forming and breaking of chemical bonds between atoms, with no change to the nuclei, and can often be described by a chemical equation. Nuclear chemistry is a sub-discipline of chemistry that involves the chemical reactions of unstable and radioactive elements where both electronic and nuclear changes can occur.
Products are the species formed from chemical reactions. During a chemical reaction reactants are transformed into products after passing through a high energy transition state. This process results in the consumption of the reactants. It can be a spontaneous reaction or mediated by catalysts which lower the energy of the transition state, and by solvents which provide the chemical environment necessary for the reaction to take place. When represented in chemical equations products are by convention drawn on the right-hand side, even in the case of reversible reactions. The properties of products such as their energies help determine several characteristics of a chemical reaction such as whether the reaction is exergonic or endergonic. Additionally the properties of a product can make it easier to extract and purify following a chemical reaction, especially if the product has a different state of matter than the reactants. Reactants are molecular materials used to create chemical reactions. The atoms aren't created or destroyed. The materials are reactive and reactants are rearranging during a chemical reaction. Here is an example of reactants: CH4 + O2. A non-example is CO2 + H2O or "energy".
In biochemistry, a lyase is an enzyme that catalyzes the breaking of various chemical bonds by means other than hydrolysis and oxidation, often forming a new double bond or a new ring structure. The reverse reaction is also possible. For example, an enzyme that catalyzed this reaction would be a lyase:
The Entner–Doudoroff pathway describes a pathway—a series of enzyme-catalyzed chemical reactions—that are active in bacterial primary metabolism, a pathway that catabolizes glucose to pyruvic acid using enzymes distinct either from those used in glycolysis or the pentose phosphate pathway. The ED pathway was first reported by Michael Doudoroff and Nathan Entner from the Bacteriology Department at the University of California, Berkeley in 1952. Recent work on the Entner–Duodoroff pathway has shown that its use is not restricted to prokaryotes as was previously thought. Specifically, there is direct evidence that Hordeum vulgare uses the Entner–Doudoroff pathway. Use of the Entner–Duodoroff pathway among other plants such as mosses and ferns is also probably widespread, based on preliminary sequencing data analysis.
Metabolism is the set of life-sustaining chemical reactions in organisms. The three main purposes of metabolism are: the conversion of food to energy to run cellular processes; the conversion of food/fuel to building blocks for proteins, lipids, nucleic acids, and some carbohydrates; and the elimination of nitrogenous wastes. These enzyme-catalyzed reactions allow organisms to grow and reproduce, maintain their structures, and respond to their environments..
In addition to the cleavage of 2-dehydro-3-deoxy-D-gluconate 6-phosphate, it is also found to naturally catalyze Schiff base formation between a lysine E-amino acid group and carbonyl compounds, decarboxylation of oxaloacetate, and exchange of solvent protons with the methyl hydrogen atoms of pyruvate.[2]
A Schiff base (named after Hugo Schiff) is a compound with the general structure R2C=NR' (R' ≠ H). They can be considered a sub-class of imines, being either secondary ketimines or secondary aldimines depending on their structure. The term is often synonymous with azomethine which refers specifically to secondary aldimines (i.e. R-CH=NR' where R' ≠ H).
Lysine (symbol Lys or K) is an α-amino acid that is used in the biosynthesis of proteins. It contains an α-amino group (which is in the protonated −NH3+ form under biological conditions), an α-carboxylic acid group (which is in the deprotonated −COO− form under biological conditions), and a side chain lysyl ((CH2)4NH2), classifying it as a basic, charged (at physiological pH), aliphatic amino acid. It is encoded by the codons, AAA and AAG. Like almost all other amino acids, the α-carbon is chiral and lysine may refer to either enantiomer or a racemic mixture of both. For the purpose of this article, lysine will refer to the biologically active enantiomer L-lysine, where the α-carbon is in the S configuration.
Nomenclature
The systematic name of this enzyme class is 2-dehydro-3-deoxy-D-gluconate-6-phosphate D-glyceraldehyde-3-phosphate-lyase (pyruvate-forming). Other names in common use include:
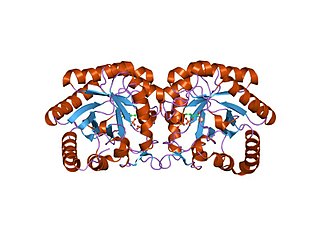
Active site structure with bound pyruvate (colors correspond to secondary structure, with cyan, magenta, and light pink referring to helices, sheets, and loops respectively)
KDPG Aldolase was recently determined to be a trimer through crystallographic three-fold symmetry, with 225 residues.[2][3] The enzyme was determined to have a molecular weight of 23,942.[4] The trimer is stabilized primarily through hydrophobic interactions. The molecule has tertiary folding similar to triosephosphate isomerase and the A-domain of pyruvate kinase, forming an eight strand α/β-barrel structure.[3][5] The α/β-barrel structure is capped on one side by the N-terminal helix. The other side, the carboxylic side, contains the active site.[6] Each subunit contains a phosphate-ion bound in position of the aldolase biding site.[7] It has been found that there are four cysteinyl groups present in each subunit, with two readily accessible and two buried in the subunit.[8]
Triose-phosphate isomerase is an enzyme that catalyzes the reversible interconversion of the triose phosphate isomers dihydroxyacetone phosphate and D-glyceraldehyde 3-phosphate.
Pyruvate kinase is the enzyme that catalyzes the final step of glycolysis. It catalyzes the transfer of a phosphate group from phosphoenolpyruvate (PEP) to adenosine diphosphate (ADP), yielding one molecule of pyruvate and one molecule of ATP. Pyruvate kinase was inappropriately named before it was recognized that it did not directly catalyze phosphorylation of phosphoenolpyruvate (PEP), which does not occur under physiological conditions. Pyruvate kinase is present in four distinct, tissue-specific isozymes in animals, each consisting of particular kinetic properties necessary to accommodate the variations in metabolic requirements of diverse tissues.
A Phosphate is a chemical derivative of phosphoric acid. The phosphate ion is an inorganic chemical, the conjugate base that can form many different salts. In organic chemistry, a phosphate, or organophosphate, is an ester of phosphoric acid. Of the various phosphoric acids and phosphates, organic phosphates are important in biochemistry and biogeochemistry, and inorganic phosphates are mined to obtain phosphorus for use in agriculture and industry. At elevated temperatures in the solid state, phosphates can condense to form pyrophosphates.
The active site contains the zwitterionic pair Glu-45/Lys-133.[9] The Lysine, which is involved in the formation of the Schiff base is coordinated by a phosphate ion and two solvent water molecules.[6][7] The first water molecule serves as a shuttle between the Glutamate and the substrate, staying bound to the enzyme throughout the catalytic cycle.[7] The second water molecule is a product of the dehydration of the carbinolamine that leads to the formation of the Schiff base.[7] It also functions as the nucleophile during hydrolysis of the enzyme-product Schiff base, leading to the release of pyruvate.[7]
Active site showing interaction of pyruvate with zwitterionic pair Glu-45/Lys-133
One of the reactions KDPG Aldolase catalyzes, as in the Entner–Doudoroff pathway, is the reversible cleavage of 2-keto-3-deoxy-6-phosphogluconate (KDPG) into pyruvate and D-glyceraldehyde-3-phosphate.[9][10] This occurs through a stereospecific retro-aldol cleavage.[7] A proton transfer between the zwitterionic pair Glu-45/Lys-133 in the active site activates Lysine to serve as the nucleophile in the first step and Glutamate to aid in the base catalysis involved in the carbon-carbon cleavage.[9] Lysine Residue 133 serves as the nucleophile and attacks the carbonyl group of 2-Keto-3-deoxy-6-phosphogluconate to form a protonated carbinolamine intermediate, also known as a Schiff base intermediate.[7][9][10] The intermediate is stabilized by hydrogen bonding with residues in the active site.[9] A three carbon residue, glyceraldehyde 3-phosphate, is cleaved off through base catalysis with a water molecule and residue Glu-45.[7][9] The pyruvate is generated through the nucleophilic attack of water on the Schiff-base to reform a ketone. Aromatic interaction with Phe-135 ensures the stereospecific addition involved in the reverse process.[9]
KDPG aldolase has also been shown to catalyze the exchange of hydrogen atoms of the methyl groups of pyruvate with protons of the solvent.[10]
KDPG aldolase mechanism in the Entner–Doudoroff pathway
Evolutionary significance
History
Arguments have been made for both the convergent and divergent evolution of α/β-barrel structured enzymes such as KDPG Aldolase, triosephosphate isomerase, and the A-domain of pyruvate kinase.
Convergent evolution can lead to geometrically similar active sites while each enzyme has a distinct backbone conformation. Convergence to a common backbone structure, as is the case here however, has not been observed, although it is argues that it might be possible for a symmetrically repetitive structure as the one observed here.[11] The similarity in the folding of the three enzymes and the exceptional symmetry commonly suggests divergent evolution from a common ancestor. The functional similarity of the enzymes remains the strongest argument for divergent evolution.[11] All three enzymes activate a C–H bond adjacent to a carbonyl group. The active sites are located at the carboxylic ends of the β strands. Such congruence is in favor of divergent evolution.
Should the divergent evolution hypothesis prevail, this would suggest the existence of a class of enzymes with unrelated amino acid sequences yet analogous symmetrical structure and folding.[11]
Directed Evolution
KDPG aldolase has limited utility due to its high specificity for its natural substrates in the cleavage of KDPG and the reverse addition of D-glyceraldehyde-3-phosphate and pyruvate.[12] In vitro evolution has allowed KDPG aldolase to be converted into a more efficient aldolase with altered substrate specificity and stereoselectivity thereby improving its utility in asymmetric synthesis.[13] Rather than modifying the recognition site, the substrate is modified by moving the active site lysine from one β strand to a neighboring one.[13][14] The evolved aldolase is capable of accepting both D- and L-glyceraldehyde in their non-phosphorylated form.[15]
Related Research Articles
Aldolase B also known as fructose-bisphosphate aldolase B or liver-type aldolase is one of three isoenzymes of the class I fructose 1,6-bisphosphate aldolase enzyme, and plays a key role in both glycolysis and gluconeogenesis. The generic fructose 1,6-bisphosphate aldolase enzyme catalyzes the reversible cleavage of fructose 1,6-bisphosphate (FBP) into glyceraldehyde 3-phosphate and dihydroxyacetone phosphate (DHAP) as well as the reversible cleavage of fructose 1-phosphate (F1P) into glyceraldehyde and dihydroxyacetone phosphate. In mammals, aldolase B is preferentially expressed in the liver, while aldolase A is expressed in muscle and erythrocytes and aldolase C is expressed in the brain. Slight differences in isozyme structure result in different activities for the two substrate molecules: FBP and fructose 1-phosphate. Aldolase B exhibits no preference and thus catalyzes both reactions, while aldolases A and C prefer FBP.
Transaldolase is an enzyme of the non-oxidative phase of the pentose phosphate pathway. In humans, transaldolase is encoded by the TALDO1 gene.
In enzymology, a phosphogluconate 2-dehydrogenase (EC 1.1.1.43) is an enzyme that catalyzes the chemical reaction
In enzymology, a phosphogluconate dehydrogenase (decarboxylating) (EC 1.1.1.44) is an enzyme that catalyzes the chemical reaction
In enzymology, a 2-dehydro-3-deoxy-D-gluconate 5-dehydrogenase (EC 1.1.1.127) is an enzyme that catalyzes the chemical reaction
In enzymology, a 2-dehydro-3-deoxy-D-gluconate 6-dehydrogenase (EC 1.1.1.126) is an enzyme that catalyzes the chemical reaction
In enzymology, a glucosaminate ammonia-lyase (EC 4.3.1.9) is an enzyme that catalyzes the chemical reaction
In enzymology, a 2-dehydro-3-deoxy-6-phosphogalactonate aldolase is an enzyme that catalyzes the chemical reaction
In enzymology, a 2-dehydro-3-deoxy-D-pentonate aldolase is an enzyme that catalyzes the chemical reaction
In enzymology, a 2-dehydro-3-deoxyglucarate aldolase is an enzyme that catalyzes the chemical reaction
In enzymology, a 2-dehydro-3-deoxy-L-pentonate aldolase is an enzyme that catalyzes the chemical reaction
In enzymology, a 5-dehydro-2-deoxyphosphogluconate aldolase is an enzyme that catalyzes the chemical reaction
In enzymology, a deoxyribose-phosphate aldolase is an enzyme that catalyzes the chemical reaction
In enzymology, a phosphogluconate dehydratase (EC 4.2.1.12) is an enzyme that catalyzes the chemical reaction
In enzymology, a 3-deoxy-8-phosphooctulonate synthase (EC 2.5.1.55) is an enzyme that catalyzes the chemical reaction
In enzymology, a 2-dehydro-3-deoxygluconokinase is an enzyme that catalyzes the chemical reaction
3-Deoxy-D-arabinoheptulosonate 7-phosphate (DAHP) synthase is the first enzyme in a series of metabolic reactions known as the shikimate pathway, which is responsible for the biosynthesis of the amino acids phenylalanine, tyrosine, and tryptophan. Since it is the first enzyme in the shikimate pathway, it controls the amount of carbon entering the pathway. Enzyme inhibition is the primary method of regulating the amount of carbon entering the pathway. Forms of this enzyme differ between organisms, but can be considered DAHP synthase based upon the reaction that is catalyzed by this enzyme.
The KDPG-aldolase RNA motif is a conserved RNA structure that was discovered by bioinformatics. KDPG-aldolase motifs are found in Enterobacteriaceae, but is not known to be bpresent in Escherichia coli.
1 2 Mavridis IM, Tulinsky A (October 1976). "The folding and quaternary structure of trimeric 2-keto-3-deoxy-6-phosphogluconic aldolase at 3.5-A resolution". Biochemistry. 15 (20): 4410–7. doi:10.1021/bi00665a010. PMID974067.
1 2 Mavridis IM, Hatada MH, Tulinsky A, Lebioda L (December 1982). "Structure of 2-keto-3-deoxy-6-phosphogluconate aldolase at 2 . 8 A resolution". J. Mol. Biol. 162 (2): 419–44. doi:10.1016/0022-2836(82)90536-8. PMID7161801.
↑ Möhler H, Decker K, Wood WA (July 1972). "Structure of 2-keto-3-deoxy-6-phosphogluconate aldolase. IV. Structural features revealed by treatment with urea and Ellman's reagent". Arch. Biochem. Biophys. 151 (1): 251–60. doi:10.1016/0003-9861(72)90495-x. PMID5044518.
1 2 3 Lebioda L, Hatada MH, Tulinsky A, Mavridis IM (December 1982). "Comparison of the folding of 2-keto-3-deoxy-6-phosphogluconate aldolase, triosephosphate isomerase and pyruvate kinase. Implications in molecular evolution". J. Mol. Biol. 162 (2): 445–58. doi:10.1016/0022-2836(82)90537-X. PMID7161802.
1 2 Fong S, Machajewski TD, Mak CC, Wong C (November 2000). "Directed evolution of D-2-keto-3-deoxy-6-phosphogluconate aldolase to new variants for the efficient synthesis of D- and L-sugars". Chem. Biol. 7 (11): 873–83. doi:10.1016/S1074-5521(00)00035-1. PMID11094340.
↑ Wymer N, Buchanan LV, Henderson D, et al. (January 2001). "Directed evolution of a new catalytic site in 2-keto-3-deoxy-6-phosphogluconate aldolase from Escherichia coli". Structure. 9 (1): 1–9. doi:10.1016/S0969-2126(00)00555-4. PMID11342129.
↑ Huisman GW, Gray D (August 2002). "Towards novel processes for the fine-chemical and pharmaceutical industries". Curr. Opin. Biotechnol. 13 (4): 352–8. doi:10.1016/S0958-1669(02)00335-X. PMID12323358.
Further reading
Meloche HP, Wood WA (1964). "Crystallization and characteristics of 2-keto-3-deoxy-6-phosphogluconic aldolase". J. Biol. Chem. 239: 3515–3518. PMID14245411.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.