Steps of the cell cycle. The restriction point occurs between the G1 and S phases of interphase. The G2-M checkpoint occurs between the G2 and M phases. The spindle checkpoint occurs during the M phase. Key cyclins associated with each phase are shown.
Cell cycle checkpoints are control mechanisms in the eukaryoticcell cycle which ensure its proper progression. Each checkpoint serves as a potential termination point along the cell cycle, during which the conditions of the cell are assessed, with progression through the various phases of the cell cycle occurring only when favorable conditions are met. There are many checkpoints in the cell cycle,[1] but the three major ones are: the G1 checkpoint, also known as the Start or restriction checkpoint or Major Checkpoint; the G2/M checkpoint; and the metaphase-to-anaphase transition, also known as the spindle checkpoint.[2] Progression through these checkpoints is largely determined by the activation of cyclin-dependent kinases by regulatory protein subunits called cyclins, different forms of which are produced at each stage of the cell cycle to control the specific events that occur therein.[3][4]
All living organisms are the products of repeated rounds of cell growth and division.[5] During this process, known as the cell cycle, a cell duplicates its contents and then divides in two. The purpose of the cell cycle is to accurately duplicate each organism's DNA and then divide the cell and its contents evenly between the two resulting cells. In eukaryotes, the cell cycle consists of four main stages: G1, during which a cell is metabolically active and continuously grows; S phase, during which DNA replication takes place; G2, during which cell growth continues and the cell synthesizes various proteins in preparation for division; and the M (mitosis) phase, during which the duplicated chromosomes (known as the sister chromatids) separate into two daughter nuclei, and the cell divides into two daughter cells, each with a full copy of DNA.[6] Compared to the eukaryotic cell cycle, the prokaryotic cell cycle (known as binary fission) is relatively simple and quick: the chromosome replicates from the origin of replication, a new membrane is assembled, and the cell wall forms a septum which divides the cell into two.[7]
As the eukaryotic cell cycle is a complex process, eukaryotes have evolved a network of regulatory proteins, known as the cell cycle control system, which monitors and dictates the progression of the cell through the cell cycle.[5] This system acts like a timer, or a clock, which sets a fixed amount of time for the cell to spend in each phase of the cell cycle, while at the same time it also responds to information received from the processes it controls. The cell cycle checkpoints play an important role in the control system by sensing defects that occur during essential processes such as DNA replication or chromosome segregation, and inducing a cell cycle arrest in response until the defects are repaired.[8] The main mechanism of action of the cell cycle checkpoints is through the regulation of the activities of a family of protein kinases known as the cyclin-dependent kinases (CDKs), which bind to different classes of regulator proteins known as cyclins, with specific cyclin-CDK complexes being formed and activated at different phases of the cell cycle. Those complexes, in turn, activate different downstream targets to promote or prevent cell cycle progression.[9]
The G1 checkpoint, also known as the restriction point in mammalian cells and the start point in yeast, is the point at which the cell becomes committed to entering the cell cycle. As the cell progresses through G1, depending on internal and external conditions, it can either delay G1, enter a quiescent state known as G0, or proceed past the restriction point.[5] DNA damage is the main indication for a cell to "restrict" and not enter the cell cycle. The decision to commit to a new round of cell division occurs when the cell activates cyclin-CDK-dependent transcription which promotes entry into S phase. This check point ensures the further process.[10]
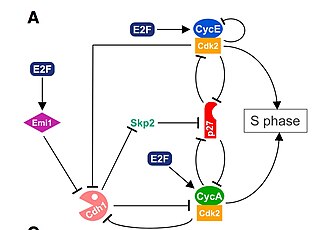
During early G1, there are three transcriptional repressors, known as pocket proteins, that bind to E2F transcription factors. The E2F gene family is a group of transcription factors that target many genes that are important for control of the cell cycle, including cyclins, CDKs, checkpoint regulators, and DNA repair proteins. Misregulation of the E2F family is often found in cancer cases, providing evidence that the E2F family is essential for the tight regulation of DNA replication and division.[10] The three pocket proteins are Retinoblastoma (Rb), p107, and p130, which bind to the E2F transcription factors to prevent progression past the G1 checkpoint.
The E2F gene family contains some proteins with activator mechanisms and some proteins with repressing mechanisms. P107 and p130 act as co-repressors for E2F 4 and E2F 5, which work to repress transcription of G1-to-S promoting factors. The third pocket protein, Rb, binds to and represses E2F 1, E2F 2, and E2F 3, which are the E2F proteins with activating abilities.[10]
Positive feedback plays an essential role in regulating the progression from G1 to S phase, particularly involving the phosphorylation of Rb by a Cyclin/CDK protein complex. Rb without a phosphate, or unphosphorylated Rb, regulates G0 cell cycle exit and differentiation. During the beginning of the G1 phase, growth factors and DNA damage signal for the rise of cyclin D levels, which then binds to Cdk4 and Cdk6 to form the CyclinD:Cdk4/6 complex.[11] This complex is known to inactivate Rb by phosphorylation. However, the details of Rb phosphorylation are quite complex and specific compared to previous knowledge about the G1checkpoint. CyclinD:Cdk4/6 places only one phosphate, or monophosphorylates, Rb at one of its fourteen accessible and unique phosphorylation sites. Each of the fourteen specific mono-phosphorylated isoforms has a differential binding preference to E2F family members, which likely adds to the diversity of cellular processes within the mammalian body.[11]
E2F 4 and E2F 5 are dependent on p107 and p130 to maintain their nuclear localization. However, Cyclin D:Cdk 4/6 also phosphorylates p107 and p130, a process which releases their bind from E2F 4 and 5 (which then escape to the cytoplasm), and allowing for E2F 1–3 to bind to the DNA and initiate transcription of Cyclin E.[10] Rb proteins maintain their mono-phosphorylated state during early G1 phase, while Cyclin E is accumulating and binding to Cdk2.
CyclinE:Cdk2 plays an additional important phosphorylation role in the G1-to-S transition. Particularly, CyclinE:Cdk2 promotes a positive feedback loop which creates an “all or nothing” switch. In many genetic control networks, positive feedback ensures that cells do not slip back and forth between cell cycle phases [12] Cyclin E:Cdk2 proceeds to phosphorylate Rb at all of its phosphorylation sites, also termed “hyper-phosphorylate”, which ensures complete inactivation of Rb. The hyper phosphorylation of Rb is considered the late G1 restriction point, after which the cell cannot go backwards in the cell cycle. At this point, E2F 1-3 proteins bind to DNA and transcribe Cyclin A and Cdc 6.[11]
Cyclin-dependent kinase inhibitor 1B (CDKN1B), also known as p27, binds to and prevents the activation of CyclinE:Cdk2 by inhibition. However, as Cyclin A accumulates and binds to Cdk2, they form a complex and inhibit p27. The G1 phase cyclin-dependent kinase works together with S phase cyclin-dependent kinase targeting p27 for degradation. In turn, this allows for full activation of Cyclin A:Cdk2, a complex which phosphorylates E2F 1-3 initiating their disassociation from the DNA promoter sites. This allows E2F 6–8 to bind to the DNA and inhibit transcription.[10] The negative feedback loop used to successfully inhibit the inhibitor, p27, is another essential process used by cells to ensure mono-directional movement and no backtrack through the cell cycle.
When DNA damage occurs, or when the cell detects any defects which necessitate it to delay or halt the cell cycle in G1, arrest occurs through several mechanisms. The rapid response involves phosphorylation events that initiate with either kinase ATM (Ataxia telangiectasia mutated) or ATR (Ataxia Telangiectasia and Rad3 related), which act as sensors, depending on the type of damage. These kinases phosphorylate and activate the effector kinases Chk2 and Chk1, respectively, which in turn phosphorylate the phosphatase Cdc25A, thus marking it for ubiquitination and degradation. As Cdc25A activates the previously mentioned cyclin E-CDK2 complex by removing inhibitory phosphates from CDK2, in the absence of Cdc25A, cyclin E-CDK2 remains inactive, and the cell remains in G1.
To maintain the arrest, another response is initiated, by which Chk2 or Chk1 phosphorylate p53, a tumor suppressor, and this stabilizes p53 by preventing it from binding Mdm2, a ubiquitin ligase which inhibits p53 by targeting it for degradation. The stable p53 then acts a transcriptional activator of several target genes, including p21, an inhibitor of the G1-to-S promoting complex cyclin E-CDK2. In addition, another mechanism by which p21 is activated is through the accumulation of p16 in response to DNA damage. p16 disrupts cyclin D-CDK4 complexes, thus causing the release of p21 from the complexes, which leads to the dephosphorylation and activation of Rb, which allows Rb to bind and inhibit E2F 1–3, thus keeping the cell from transitioning to S phase.[13] Recently, some aspects of this model have been disputed.[14]
Mitotic Cyclin Concentration shows hysteresis and bistability relative to Cdk1 Activation
Following DNA replication in S phase, the cell undergoes a growth phase known as G2. During this time, necessary mitotic proteins are produced and the cell is once more subjected to regulatory mechanisms to ensure proper status for entry into the proliferative Mitotic (M) phase. Multiple mechanistic checkpoints are involved in this transition from G2 to M, with a common uniting factor of cyclin-Cdk activity.
Although variations in requisite cyclin-Cdk complexes exist across organisms, the necessity of the kinase activity is conserved and typically focuses on a single pairing. In fission yeast three different forms of mitotic cyclin exist, and six in budding yeast, yet the primary cyclin utilized is cyclin B.[15] Cyclin B will serve as reference for discussion of the G2/M checkpoint transition.
Similar to S Phase, G2 experiences a DNA damage checkpoint. The cell is once more examined for sites of DNA damage or incomplete replication, and the kinases ATR and ATM are recruited to damage sites. Activation of Chk1 and Chk2 also transpire, as well as p53 activation, to induce cell cycle arrest and halt progression into mitosis. An additional component of S phase, the Pre-Replicative Complex, must be inactivated via cyclin B-Cdk1 phosphorylation.[16]
As these previous checkpoints are assessed, G2 protein accumulation serves to activate cyclin B-Cdk1 activity via multiple mechanisms. CyclinA-Cdk2 activates Cdc25, an activator of cyclin B-Cdk1, which then deactivates the cyclin B-Cdk1 inhibitor, Wee1. This results in a positive feedback loop, significantly increasing cyclin B expression and Cdk1 activation. As the cell progresses through G2 and reaches the G2/M transition, the kinase Plk1 phosphorylates Wee1, which targets Wee1 for degradation via the SCF ubiquitin ligase complex.[17] An additional function of Plk1 is to activate Cdc25 through phosphorylation. The compound effect of Wee1 degradation and Cdc25 activation is the net removal of inhibitory phosphorylation from cdc2, which activates cdc2. Plk1 is activated at the G2/M transition by the Aurora A and Bora, which accumulate during G2 and form an activation complex. The Plk1-Cdc2-cdc25 complex then initiates a positive feedback loop which serves to further activate Cdc2, and in conjunction with an increase in cyclin B levels during G2, the resulting cdc2-cyclin B complexes then activate downstream targets which promote entry into mitosis.[18] The resultant Cdk1 activity also activates expression of Mem1-Fkh, a G2/M transition gene.[19] The rapid surge in cyclin B-Cdk1 activity is necessary, as M phase initiation is an all-or-nothing event engaging in hysteresis. Hysteresis of Cdk1 activity via cyclin B drives M phase entry by establishing a minimum threshold of cyclin B concentration. This exists at a level higher than the minimum needed for the continuation of M phase after entry, acting to safeguard the all-or-nothing event. This entry concentration is further increased in the case of incomplete DNA replication, adding another regulatory mechanism at the G2/M transition point.[20] The presence of hysteresis allows for M phase entry to be highly regulated as a function of cyclin B-Cdk1 activity.
The mechanisms by which mitotic entry is prevented in response to DNA damage are similar to those in the G1/S checkpoint. DNA damage triggers the activation of the aforementioned ATM/ATR pathway, in which ATM/ATR phosphorylate and activate the Chk1/Chk2 checkpoint kinases. Chk1/2 phosphorylate cdc25 which, in addition to being inhibited, is also sequestered in the cytoplasm by the 14-3-3 proteins. 14-3-3 are upregulated by p53, which, as previously mentioned, is activated by Chk1 and ATM/ATR. p53 also transactivates p21, and both p21 and the 14-3-3 in turn inhibit cyclin B-cdc2 complexes through the phosphorylation and cytoplasmic sequestering of cdc2. In addition, the inactivation of cdc25 results in its inability to dephosphorylate and activate cdc2.[21][22] Finally, another mechanism of damage response is through the negative regulation of Plk1 by ATM/ATR, which in turn results in the stabilization of Wee1 and Myt1, which can then phosphorylate and inhibit cdc2, thus keeping the cell arrested in G2 until the damage is fixed.[23]
G2–M transition in Xenopus oocytes
At the end of G2, the cell transitions into mitosis, where the nucleus divides. The G2 to M transition is dramatic; there is an all-or-nothing effect, and the transition is irreversible. This is advantageous to the cell because entering mitosis is a critical step in the life cycle of a cell. If it does not fully commit, the cell would run into many issues with partially dividing, ultimately likely leading to the cell's death.
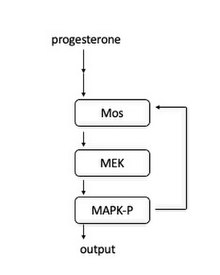
In frog oocytes, the signal cascade is induced when progesterone binds to a membrane bound receptor. Downstream, Mos is activated. Mos then phosphorylates MEK1, which phosphorylates MAPK. MAPK serves two roles: activating the Cyclin B-Cdk1 complex to initiate entrance into mitosis and activating Mos . The activation of Mos leads to a positive feedback loop and therefore acts as “toggle switch” to create the all-or-nothing entrance into mitosis.
Schematic of the MAPK signaling cascade.
This feedback loop was first found by showing that MAPK-P (phosphorylated MAPK) concentrations increased in response to increasing levels of progesterone.[24] At the single cell level, each cell either had entirely phosphorylated MAPK or no phosphorylated MAPK, confirming that it acts as a switch-like mechanism in each cell. It was additionally shown that blocking Mos protein synthesis makes the MAPK-P responses more graded, showing that Mos protein synthesis is necessary for the all-or-none character of MAPK activation.[25]
Bistability
This process can be understood using unstability. Using the graph shown to the right, the Mos synthesis rate shifts as more progesterone is added. With each curve, there are stable fixed points and unstable fixed points. At the unstable fixed points, the system will push toward either one of the stable fixed points. So, the system can either be in the “on” state or the “off” state, not in between. When the progesterone level is high enough, the Mos curve is shifted higher and ultimately intersects the degradation line at only one point, so there is only one stable “on” state, indicating the entrance into mitosis.
The irreversibility we see in the Mitosis transition point comes from having high enough levels of progesterone in the cell. At high enough levels of progesterone, the system is monostable as a result of the positive feedback loop between Mapk and Mos. The point at which the system switches from bistable to monostable is called the saddle node bifurcation.
So, we can understand the all-or-nothing, irreversible response of the mitotic transition with a mathematical model of the molecular regulators as a bistable system that depends on the existence of positive feedback. The “off-state” is annihilated by a high enough level of progesterone and once the cell gets pushed past the off-state, it is then stuck in the on-state.
Hysteresis and the Novak–Tyson model
Coming from this bi-stable model, we can understand the mitotic transition as relying on hysteresis to drive it. Hysteresis is defined as the dependence of the state of a system on its history. The Novak–Tyson model is a mathematical model of cell cycle progression that predicts that irreversible transitions entering and exiting mitosis are driven by hysteresis. The model has three basic predictions that should hold true in cycling oocyte extracts whose cell cycle progression is dependent on hysteresis:[26]
The concentration of cyclin B necessary to enter mitosis is higher than the concentration needed to hold a mitotic extract in mitosis.
Unreplicated DNA raises the level of cyclin necessary for Cdc2 activation and therefore entrance into mitosis.
There is a decrease in the rate of Cdc2 activation at concentrations of cyclin B just above the activation threshold.
Sha et al. did experiments in Xenopus laevis egg extracts in 2003 to demonstrate this hysteretic nature.[27] Using cycling extracts, they observed that the activation threshold for Δcyclin B is between 32 and 42 nM whereas the inactivation threshold is between 16 and 24 nM Δcyclin B. Therefore, these experiments confirmed the bistability of this system and the importance of hysteresis in this cell cycle transition. At the intermediate cyclin B concentrations, either the interphase or mitotic state of the cell is possible.
Replication stress response
Since entering mitosis is a large and costly commitment for the cell, it is logical that systems would be in place to prevent premature entrance into this step. It has been shown that mistakes in previous steps, such as having unreplicated sections of DNA blocks progression in the cell cycle.[28] The Novak–Tyson model predicts this occurs via raising the level of cyclin B necessary for entrance into mitosis.[26]
Sha et al. investigated whether this was true in Xenopus egg extracts. They used aphidicolin (APH) to inhibit DNA polymerase and prevent DNA replication. When treated with Cyclin B in interphase, the threshold of activation increased to between 80 and 100 nM, as predicted by the Novak–Tyson model.[27] So, these experiments confirm that the stress of unreplicated DNA in the cell affect the hysteresis loop and result in a much higher cyclin B threshold to enter into mitosis.
Metaphase checkpoint
Activation of the mitotic spindle checkpoint blocks entry into anaphase.
The mitotic spindle checkpoint occurs at the point in metaphase where all the chromosomes should/have aligned at the mitotic plate and be under bipolar tension. The tension created by this bipolar attachment is what is sensed, which initiates the anaphase entry. To do this, the sensing mechanism ensures that the anaphase-promoting complex (APC/C) is no longer inhibited, which is now free to degrade cyclin B, which harbors a D-box (destruction box), and to break down securin.[29] The latter is a protein whose function is to inhibit separase, which in turn cuts the cohesins, the protein composite responsible for cohesion of sister chromatids.[30] Once this inhibitory protein is degraded via ubiquitination and subsequent proteolysis, separase then causes sister chromatid separation.[31] After the cell has split into its two daughter cells, the cell enters G1.
Cancer
DNA repair processes and cell cycle checkpoints have been intimately linked with cancer due to their functions regulating genome stability and cell progression, respectively. The precise molecular mechanisms that connect dysfunctions in these pathways to the onset of particular cancers are not well understood in most cases.[32] The loss of ATM has been shown to precede lymphoma development presumably due to excessive homologous recombination, leading to high genomic instability.[33] Disruption of Chk1 in mice led significant misregulation of cell cycle checkpoints, an accumulation of DNA damage, and an increased incidence of tumorigenesis.[34] Single mutant inheritance of BRCA1 or BRCA2 predisposes females toward breast and ovarian cancers.[35] BRCA1 is known to be required for S and G2/M transitions, and is involved in the cellular response to DNA damage. BRCA2 is believed to be involved in homologous recombination and regulating the S-phase checkpoint, and mutations of deficiencies in BRCA2 are strongly linked to tumorigenesis.[36]
The cell cycle, or cell-division cycle, is the sequential series of events that take place in a cell that causes it to divide into two daughter cells. These events include the growth of the cell, duplication of its DNA and some of its organelles, and subsequently the partitioning of its cytoplasm, chromosomes and other components into two daughter cells in a process called cell division.
Cyclins are proteins that control the progression of a cell through the cell cycle by activating cyclin-dependent kinases (CDK).
Cyclin-dependent kinases (CDKs) are a predominant group of serine/threonine protein kinases involved in the regulation of the cell cycle and its progression, ensuring the integrity and functionality of cellular machinery. These regulatory enzymes play a crucial role in the regulation of eukaryotic cell cycle and transcription, as well as DNA repair, metabolism, and epigenetic regulation, in response to several extracellular and intracellular signals. They are present in all known eukaryotes, and their regulatory function in the cell cycle has been evolutionarily conserved. The catalytic activities of CDKs are regulated by interactions with CDK inhibitors (CKIs) and regulatory subunits known as cyclins. Cyclins have no enzymatic activity themselves, but they become active once they bind to CDKs. Without cyclin, CDK is less active than in the cyclin-CDK heterodimer complex. CDKs phosphorylate proteins on serine (S) or threonine (T) residues. The specificity of CDKs for their substrates is defined by the S/T-P-X-K/R sequence, where S/T is the phosphorylation site, P is proline, X is any amino acid, and the sequence ends with lysine (K) or arginine (R). This motif ensures CDKs accurately target and modify proteins, crucial for regulating cell cycle and other functions. Deregulation of the CDK activity is linked to various pathologies, including cancer, neurodegenerative diseases, and stroke.
A cyclin-dependent kinase complex is a protein complex formed by the association of an inactive catalytic subunit of a protein kinase, cyclin-dependent kinase (CDK), with a regulatory subunit, cyclin. Once cyclin-dependent kinases bind to cyclin, the formed complex is in an activated state. Substrate specificity of the activated complex is mainly established by the associated cyclin within the complex. Activity of CDKCs is controlled by phosphorylation of target proteins, as well as binding of inhibitory proteins.
Maturation-promoting factor (abbreviated MPF, also called mitosis-promoting factor or M-Phase-promoting factor) is the cyclin–Cdk complex that was discovered first in frog eggs. It stimulates the mitotic and meiotic phases of the cell cycle. MPF promotes the entrance into mitosis (the M phase) from the G2 phase by phosphorylating multiple proteins needed during mitosis. MPF is activated at the end of G2 by a phosphatase, which removes an inhibitory phosphate group added earlier.
S phase (Synthesis phase) is the phase of the cell cycle in which DNA is replicated, occurring between G1 phase and G2 phase. Since accurate duplication of the genome is critical to successful cell division, the processes that occur during S-phase are tightly regulated and widely conserved.
G2 phase, Gap 2 phase, or Growth 2 phase, is the third subphase of interphase in the cell cycle directly preceding mitosis. It follows the successful completion of S phase, during which the cell’s DNA is replicated. G2 phase ends with the onset of prophase, the first phase of mitosis in which the cell’s chromatin condenses into chromosomes.
The restriction point (R), also known as the Start or G1/S checkpoint, is a cell cycle checkpoint in the G1 phase of the animal cell cycle at which the cell becomes "committed" to the cell cycle, and after which extracellular signals are no longer required to stimulate proliferation. The defining biochemical feature of the restriction point is the activation of G1/S- and S-phase cyclin-CDK complexes, which in turn phosphorylate proteins that initiate DNA replication, centrosome duplication, and other early cell cycle events. It is one of three main cell cycle checkpoints, the other two being the G2-M DNA damage checkpoint and the spindle checkpoint.
The G1/S transition is a stage in the cell cycle at the boundary between the G1 phase, in which the cell grows, and the S phase, during which DNA is replicated. It is governed by cell cycle checkpoints to ensure cell cycle integrity and the subsequent S phase can pause in response to improperly or partially replicated DNA. During this transition the cell makes decisions to become quiescent, differentiate, make DNA repairs, or proliferate based on environmental cues and molecular signaling inputs. The G1/S transition occurs late in G1 and the absence or improper application of this highly regulated checkpoint can lead to cellular transformation and disease states such as cancer.
Cyclin A is a member of the cyclin family, a group of proteins that function in regulating progression through the cell cycle. The stages that a cell passes through that culminate in its division and replication are collectively known as the cell cycle Since the successful division and replication of a cell is essential for its survival, the cell cycle is tightly regulated by several components to ensure the efficient and error-free progression through the cell cycle. One such regulatory component is cyclin A which plays a role in the regulation of two different cell cycle stages.
Cyclin-dependent kinase 2, also known as cell division protein kinase 2, or Cdk2, is an enzyme that in humans is encoded by the CDK2 gene. The protein encoded by this gene is a member of the cyclin-dependent kinase family of Ser/Thr protein kinases. This protein kinase is highly similar to the gene products of S. cerevisiae cdc28, and S. pombe cdc2, also known as Cdk1 in humans. It is a catalytic subunit of the cyclin-dependent kinase complex, whose activity is restricted to the G1-S phase of the cell cycle, where cells make proteins necessary for mitosis and replicate their DNA. This protein associates with and is regulated by the regulatory subunits of the complex including cyclin E or A. Cyclin E binds G1 phase Cdk2, which is required for the transition from G1 to S phase while binding with Cyclin A is required to progress through the S phase. Its activity is also regulated by phosphorylation. Multiple alternatively spliced variants and multiple transcription initiation sites of this gene have been reported. The role of this protein in G1-S transition has been recently questioned as cells lacking Cdk2 are reported to have no problem during this transition.
Cyclin-dependent kinase 1 also known as CDK1 or cell division cycle protein 2 homolog is a highly conserved protein that functions as a serine/threonine protein kinase, and is a key player in cell cycle regulation. It has been highly studied in the budding yeast S. cerevisiae, and the fission yeast S. pombe, where it is encoded by genes cdc28 and cdc2, respectively. With its cyclin partners, Cdk1 forms complexes that phosphorylate a variety of target substrates ; phosphorylation of these proteins leads to cell cycle progression.
G2/mitotic-specific cyclin-B1 is a protein that in humans is encoded by the CCNB1 gene.
Cyclin-A2 is a protein that in humans is encoded by the CCNA2 gene. It is one of the two types of cyclin A: cyclin A1 is expressed during meiosis and embryogenesis while cyclin A2 is expressed in the mitotic division of somatic cells.
Membrane-associated tyrosine- and threonine-specific cdc2-inhibitory kinase also known as Myt1 kinase is an enzyme that in humans is encoded by the PKMYT1 gene.
Wee1 is a nuclear kinase belonging to the Ser/Thr family of protein kinases in the fission yeast Schizosaccharomyces pombe. Wee1 has a molecular mass of 96 kDa and is a key regulator of cell cycle progression. It influences cell size by inhibiting the entry into mitosis, through inhibiting Cdk1. Wee1 has homologues in many other organisms, including mammals.
A series of biochemical switches control transitions between and within the various phases of the cell cycle. The cell cycle is a series of complex, ordered, sequential events that control how a single cell divides into two cells, and involves several different phases. The phases include the G1 and G2 phases, DNA replication or S phase, and the actual process of cell division, mitosis or M phase. During the M phase, the chromosomes separate and cytokinesis occurs.
The G2-M DNA damage checkpoint is an important cell cycle checkpoint in eukaryotic organisms that ensures that cells don't initiate mitosis until damaged or incompletely replicated DNA is sufficiently repaired. Cells with a defective G2-M checkpoint will undergo apoptosis or death after cell division if they enter the M phase before repairing their DNA. The defining biochemical feature of this checkpoint is the activation of M-phase cyclin-CDK complexes, which phosphorylate proteins that promote spindle assembly and bring the cell to metaphase.
The Neuronal cell cycle represents the life cycle of the biological cell, its creation, reproduction and eventual death. The process by which cells divide into two daughter cells is called mitosis. Once these cells are formed they enter G1, the phase in which many of the proteins needed to replicate DNA are made. After G1, the cells enter S phase during which the DNA is replicated. After S, the cell will enter G2 where the proteins required for mitosis to occur are synthesized. Unlike most cell types however, neurons are generally considered incapable of proliferating once they are differentiated, as they are in the adult nervous system. Nevertheless, it remains plausible that neurons may re-enter the cell cycle under certain circumstances. Sympathetic and cortical neurons, for example, try to reactivate the cell cycle when subjected to acute insults such as DNA damage, oxidative stress, and excitotoxicity. This process is referred to as “abortive cell cycle re-entry” because the cells usually die in the G1/S checkpoint before DNA has been replicated.
The Cyclin E/Cdk2 complex is a structure composed of two proteins, cyclin E and cyclin-dependent kinase 2 (Cdk2). Similar to other cyclin/Cdk complexes, the cyclin E/Cdk2 dimer plays a crucial role in regulating the cell cycle, with this specific complex peaking in activity during the G1/S transition. Once the cyclin and Cdk subunits join together, the complex gets activated, allowing it to phosphorylate and bind to downstream proteins to ultimately promote cell cycle progression. Although cyclin E can bind to other Cdk proteins, its primary binding partner is Cdk2, and the majority of cyclin E activity occurs when it exists as the cyclin E/Cdk2 complex.
↑ Bartek J, Lukas J (December 2001). "Mammalian G1- and S-phase checkpoints in response to DNA damage". Current Opinion in Cell Biology. 13 (6): 738–47. doi:10.1016/S0955-0674(00)00280-5. PMID11698191.
↑ Löbrich M, Jeggo PA (November 2007). "The impact of a negligent G2/M checkpoint on genomic instability and cancer induction". Nature Reviews. Cancer. 7 (11): 861–9. doi:10.1038/nrc2248. PMID17943134. S2CID30207932.
↑ Peters JM (December 1998). "SCF and APC: the Yin and Yang of cell cycle regulated proteolysis". Current Opinion in Cell Biology. 10 (6): 759–68. doi:10.1016/S0955-0674(98)80119-1. PMID9914180.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.