Function of preprophase in the cell cycle
Plant cells are fixed with regards to their neighbor cells within the tissues they are growing in. In contrast to animals where certain cells can migrate within the embryo to form new tissues, the seedlings of higher plants grow entirely based on the orientation of cell division and subsequent elongation and differentiation of cells within their cell walls. Therefore, the accurate control of cell division planes and placement of the future cell wall in plant cells is crucial for the correct architecture of plant tissues and organs.
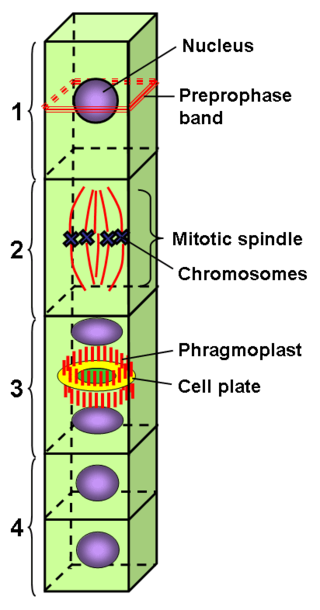
The preprophase stage of somatic plant cell mitosis serves to establish the precise location of the division plane and future cell wall before the cell enters prophase. This is achieved through the formation of a transient microtubule structure, the preprophase band, and a so far unknown mechanism by which the cell is able to "memorize" the position of the preprophase band to guide the new cell wall growing during cytokinesis to the correct location. In gametophyte tissues during the reproductive phase of the plant life cycle, cell division planes may be established without the use of a preprophase band. [2]
In highly vacuolated plant cells, preprophase may be preceded by the formation of a phragmosome. The function of the phragmosome is to suspend the cell nucleus in the center of the cell in preparation for mitosis. If a phragmosome is visible, the preprophase band will appear at its outer edge.

Mitosis is a part of the cell cycle in which replicated chromosomes are separated into two new nuclei. Cell division by mitosis is an equational division which gives rise to genetically identical cells in which the total number of chromosomes is maintained. Mitosis is preceded by the S phase of interphase and is followed by telophase and cytokinesis, which divide the cytoplasm, organelles, and cell membrane of one cell into two new cells containing roughly equal shares of these cellular components. The different stages of mitosis altogether define the mitotic phase of a cell cycle—the division of the mother cell into two daughter cells genetically identical to each other.

Cell division is the process by which a parent cell divides into two daughter cells. Cell division usually occurs as part of a larger cell cycle in which the cell grows and replicates its chromosome(s) before dividing. In eukaryotes, there are two distinct types of cell division: a vegetative division (mitosis), producing daughter cells genetically identical to the parent cell, and a cell division that produces haploid gametes for sexual reproduction (meiosis), reducing the number of chromosomes from two of each type in the diploid parent cell to one of each type in the daughter cells. Mitosis is a part of the cell cycle, in which, replicated chromosomes are separated into two new nuclei. Cell division gives rise to genetically identical cells in which the total number of chromosomes is maintained. In general, mitosis is preceded by the S stage of interphase and is followed by telophase and cytokinesis; which divides the cytoplasm, organelles, and cell membrane of one cell into two new cells containing roughly equal shares of these cellular components. The different stages of mitosis all together define the M phase of an animal cell cycle—the division of the mother cell into two genetically identical daughter cells. To ensure proper progression through the cell cycle, DNA damage is detected and repaired at various checkpoints throughout the cycle. These checkpoints can halt progression through the cell cycle by inhibiting certain cyclin-CDK complexes. Meiosis undergoes two divisions resulting in four haploid daughter cells. Homologous chromosomes are separated in the first division of meiosis, such that each daughter cell has one copy of each chromosome. These chromosomes have already been replicated and have two sister chromatids which are then separated during the second division of meiosis. Both of these cell division cycles are used in the process of sexual reproduction at some point in their life cycle. Both are believed to be present in the last eukaryotic common ancestor.
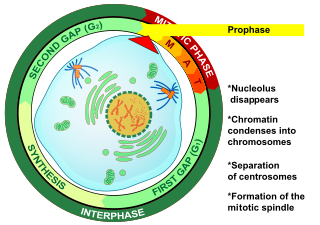
Prophase is the first stage of cell division in both mitosis and meiosis. Beginning after interphase, DNA has already been replicated when the cell enters prophase. The main occurrences in prophase are the condensation of the chromatin reticulum and the disappearance of the nucleolus.
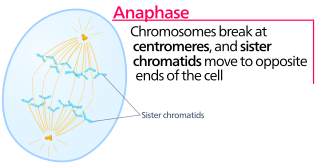
Anaphase is the stage of mitosis after the process of metaphase, when replicated chromosomes are split and the newly-copied chromosomes are moved to opposite poles of the cell. Chromosomes also reach their overall maximum condensation in late anaphase, to help chromosome segregation and the re-formation of the nucleus.

Cytokinesis is the part of the cell division process and part of mitosis during which the cytoplasm of a single eukaryotic cell divides into two daughter cells. Cytoplasmic division begins during or after the late stages of nuclear division in mitosis and meiosis. During cytokinesis the spindle apparatus partitions and transports duplicated chromatids into the cytoplasm of the separating daughter cells. It thereby ensures that chromosome number and complement are maintained from one generation to the next and that, except in special cases, the daughter cells will be functional copies of the parent cell. After the completion of the telophase and cytokinesis, each daughter cell enters the interphase of the cell cycle.

In cell biology, the spindle apparatus is the cytoskeletal structure of eukaryotic cells that forms during cell division to separate sister chromatids between daughter cells. It is referred to as the mitotic spindle during mitosis, a process that produces genetically identical daughter cells, or the meiotic spindle during meiosis, a process that produces gametes with half the number of chromosomes of the parent cell.

In cell biology, the cleavage furrow is the indentation of the cell's surface that begins the progression of cleavage, by which animal and some algal cells undergo cytokinesis, the final splitting of the membrane, in the process of cell division. The same proteins responsible for muscle contraction, actin and myosin, begin the process of forming the cleavage furrow, creating an actomyosin ring. Other cytoskeletal proteins and actin binding proteins are involved in the procedure.

Telophase is the final stage in both meiosis and mitosis in a eukaryotic cell. During telophase, the effects of prophase and prometaphase are reversed. As chromosomes reach the cell poles, a nuclear envelope is re-assembled around each set of chromatids, the nucleoli reappear, and chromosomes begin to decondense back into the expanded chromatin that is present during interphase. The mitotic spindle is disassembled and remaining spindle microtubules are depolymerized. Telophase accounts for approximately 2% of the cell cycle's duration.
The microtubule-organizing center (MTOC) is a structure found in eukaryotic cells from which microtubules emerge. MTOCs have two main functions: the organization of eukaryotic flagella and cilia and the organization of the mitotic and meiotic spindle apparatus, which separate the chromosomes during cell division. The MTOC is a major site of microtubule nucleation and can be visualized in cells by immunohistochemical detection of γ-tubulin. The morphological characteristics of MTOCs vary between the different phyla and kingdoms. In animals, the two most important types of MTOCs are 1) the basal bodies associated with cilia and flagella and 2) the centrosome associated with spindle formation.

Prometaphase is the phase of mitosis following prophase and preceding metaphase in eukaryotic somatic cells. In prometaphase, the nuclear membrane breaks apart into numerous "membrane vesicles," and the chromosomes inside form protein structures called kinetochores. Kinetochore microtubules emerging from the centrosomes at the poles (ends) of the spindle reach the chromosomes and attach to the kinetochores, throwing the chromosomes into agitated motion. Other spindle microtubules make contact with microtubules coming from the opposite pole. Forces exerted by protein "motors" associated with spindle microtubules move the chromosomes toward the centre of the cell.
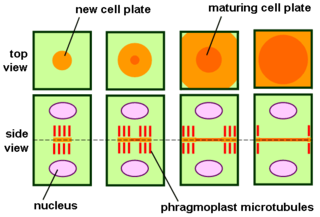
The phragmoplast is a plant cell specific structure that forms during late cytokinesis. It serves as a scaffold for cell plate assembly and subsequent formation of a new cell wall separating the two daughter cells. The phragmoplast can only be observed in Phragmoplastophyta, a clade that includes the Coleochaetophyceae, Zygnematophyceae, Mesotaeniaceae, and Embryophyta. Some algae use another type of microtubule array, a phycoplast, during cytokinesis.

Motor proteins are a class of molecular motors that can move along the cytoskeleton of cells. They convert chemical energy into mechanical work by the hydrolysis of ATP. Flagellar rotation, however, is powered by a proton pump.
The cell cortex, also known as the actin cortex, cortical cytoskeleton or actomyosin cortex, is a specialized layer of cytoplasmic proteins on the inner face of the cell membrane. It functions as a modulator of membrane behavior and cell surface properties. In most eukaryotic cells lacking a cell wall, the cortex is an actin-rich network consisting of F-actin filaments, myosin motors, and actin-binding proteins. The actomyosin cortex is attached to the cell membrane via membrane-anchoring proteins called ERM proteins that plays a central role in cell shape control. The protein constituents of the cortex undergo rapid turnover, making the cortex both mechanically rigid and highly plastic, two properties essential to its function. In most cases, the cortex is in the range of 100 to 1000 nanometers thick.

In biology, a protein filament is a long chain of protein monomers, such as those found in hair, muscle, or in flagella. Protein filaments form together to make the cytoskeleton of the cell. They are often bundled together to provide support, strength, and rigidity to the cell. When the filaments are packed up together, they are able to form three different cellular parts. The three major classes of protein filaments that make up the cytoskeleton include: actin filaments, microtubules and intermediate filaments.
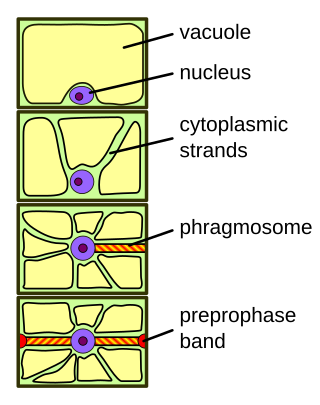
The phragmosome is a sheet of cytoplasm forming in highly vacuolated plant cells in preparation for mitosis. In contrast to animal cells, plant cells often contain large central vacuoles occupying up to 90% of the total cell volume and pushing the nucleus against the cell wall. In order for mitosis to occur, the nucleus has to move into the center of the cell. This happens during G2 phase of the cell cycle.
The preprophase band is a microtubule array found in plant cells that are about to undergo cell division and enter the preprophase stage of the plant cell cycle. Besides the phragmosome, it is the first microscopically visible sign that a plant cell is about to enter mitosis. The preprophase band was first observed and described by Jeremy Pickett-Heaps and Donald Northcote at Cambridge University in 1966.
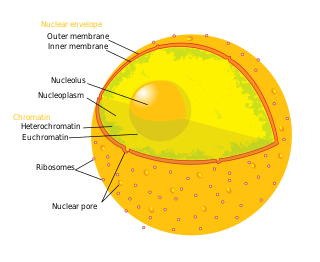
The nuclear envelope, also known as the nuclear membrane, is made up of two lipid bilayer membranes that in eukaryotic cells surround the nucleus, which encloses the genetic material.

Dynactin is a 23 subunit protein complex that acts as a co-factor for the microtubule motor cytoplasmic dynein-1. It is built around a short filament of actin related protein-1 (Arp1).

An aster is a cellular structure shaped like a star, consisting of a centrosome and its associated microtubules during the early stages of mitosis in an animal cell. Asters do not form during mitosis in plants. Astral rays, composed of microtubules, radiate from the centrosphere and look like a cloud. Astral rays are one variant of microtubule which comes out of the centrosome; others include kinetochore microtubules and polar microtubules.

Peter Klock Hepler HonFRMS is the Constantine J. Gilgut and Ray Ethan Torrey Professor Emeritus in the Biology Department of the University of Massachusetts at Amherst who is notable for his work on elucidating the roles of calcium, membranes and the cytoskeleton in plant cell development and cell motility.
